Identification of Single Nucleotide Polymorphisms (SNPs) of the Bovine Growth Hormone (bGH) Gene Associated with Growth and Carcass Traits in Hanwoo
Article information
Abstract
The purpose of this study was to find any association of the bovine growth hormone (bGH) gene with growth and carcass quality traits in Korean native cattle, Hanwoo. Genomic DNA was extracted from 21 Hanwoo individuals, and the 47 to 2,528 bp region of the bGH 2,856 bp (GenBank accession number M57764) including the promoter and the five exons was sequenced. A total of ten bGH SNPs were confirmed, including four (253 C>T, 303 C>T, 502 C>T, and 559 G>A) in the promoter, one (679 C>T) in exon 1, one (1,692 T>C) in intron 3, and four (2141 C>G, 2258 C>T, 2277 C>T, and 2291 A>C) in exon 5. The ten bGH SNPs were genotyped for a sample of 242 Hanwoo steers and association tests were performed to find any significant SNP that was correlated with growth and carcass quality. Of the SNPs, the 303 C>T SNP in the promoter region was significantly associated with 6-month-old weight, the 559 G>A SNP with longissimus dorsi muscle area, the 2141 C>G SNP in exon 5 with daily weight gain, and the 2258 C>T SNP with daily weight gain and carcass weight (p<0.05). The significant SNPs need to be verified in other Hanwoo populations before considering implementation of marker-assisted selection for genetic improvement of growth and carcass quality in Hanwoo.
INTRODUCTION
The bovine growth hormone (bGH) gene is located in q22 of bovine chromosome 19 and includes five exons with 217 amino acids (Santome et al., 1971; Wallis, 1973; Miller et al., 1980). The gene was sequenced by Gordon et al. (1983) and Hediger et al. (1990). Its product, growth hormone, is secreted in somatotropic or acidophilic cells of the anterior pituitary gland of mammals. The hormone regulates expression of many genes including one encoding insulin-like growth factor I (IGF-I), and influences growth rate, body composition, health, and milk production (Woychik et al., 1982; Gordon et al., 1983; Sumantran et al., 1992; Ho and Hoffman, 1993; Lincoln et al., 1995; Ge et al., 2013). In general, GH binds to a GH receptor and forms a dimer. Janus kinase and mitogen-activated protein kinase are involved in GH activity (Herrington et al., 2000). Recently, studies of the bGH gene have focused on single nucleotide polymorphisms (SNPs) within the gene and how the polymorphisms influence production traits such as milk production, growth, or carcass traits in cattle.
Roth et al. (1990) found a SNP in the bGH promoter region, ‘253’ SNP near the binding site of polyoma virus enhancer A binding protein 3 (PEA3) transcription factor. Theill and Karin (1993) identified ‘303’ SNP in the first nucleotide of biding site of another transcription factor, thyroid hormone response element (TRE). The 303 SNP was confirmed in eight cattle breeds (Hecht and Geldermann, 1996). Ge et al. (2013) analyzed effects of the bGH SNPs on growth traits and concentrations of IGF-I in Angus, but did not find significant evidence that the SNPs were associated with the traits. However, Kim et al. (2004) reported a bGH SNP, ‘-120’ SNP in promoter region of the gene, which was associated with 3-month weight and carcass weight in Hanwoo.
There are also many reports that bGH SNPs, especially ones in exon 5, influenced milk production in Holstein and economically important traits in beef cattle (Eppard et al., 1992; Zhang et al., 1992, 1993; Lee et al., 1993; Lucy et al., 1993; Schlee et al., 1994; Yao et al., 1996). The 2141 C>G SNP encoding the 127th amino acid of bGH involves a change from leucine (CTG) to valine (GTG) (Zhang et al., 1992, 1993; Lucy et al., 1993). For the SNP, Eppard et al. (1992) found that the GTG increased milk yields in Holstein. However, Lee et al. (1993) and Lucy et al. (1993) reported that the mutation to valine decreased milk yield in cattle. Schlee et al. (1994) demonstrated that German black and white bulls with homozygous leucine had higher plasma levels than the ones with heterozygous leucine. Yao et al. (1996) reported that the 2291 A>C bGH SNP was significantly associated with milk yield, fat, and protein content in Holstein cattle. Recently, Ardiyanti et al. (2012) reported association of bGH to fatty acid components in Japanese black cattle.
Yoon et al. (2003) found a 2258 C>T SNP of bGH in Hanwoo, causing replacement of arginine (CGG) with tryptophan (TGG). Chikuni et al. (1994) also reported a 2277 C>T SNP in bGH in Japanese cattle.
The objective of this study was to find any association of bGH SNPs with growth and carcass traits in Hanwoo.
MATERIALS AND METHODS
Animals and phenotype data
A sample of 242 Hanwoo steers from 25 sires was collected from the Korea Animal Improvement Association. All the steers were under the progeny-testing program to select Hanwoo proven sires in the National Livestock Research Institute (NLRI), Korea. The steers were raised under tightly controlled conditions of the feeding program in the Daekwanryeong and Namwon branches of NLRI. The animals were born between the spring of 1998 and the fall of 2002, castrated at 6 months of age, and raised in groups of four animals per pen (4 m×8 m). After 6 months of age, the steers were fed concentrates consisting of 15% crude protein (CP)/71% totally digestible nutrients (TDN) for a period of 60 to 90 d, 15% CP/71% TDN for a period of 180 d, and 13% CP/72% TDN for a period of 90 to 120 d. The steers had access to roughage and fresh water ad libitum throughout the entire period. All steers were slaughtered approximately at 24 months of age. Live weight of each steer was measured before slaughter using electronic scales. Following a 24-h chilling, cold carcass weight was also measured.
Growth traits included weights of six-month (WT6), 12-month (WT12), 18-month (WT18), and 24-month (WT24) of age. Average daily gain (ADG) was also measured. Carcass quality traits included carcass weight (CWT), backfat thickness (BF), longissimus dorsi (eye) muscle area (EMA), and marbling scores (MS). According to the protocols of Korean Animal Product Grade System of Korean Institute for Animal Products Quality Evaluation, BF (mm) was measured at the 2/3 point of backfat that was located toward abdomen along the right side of the eye muscle cross-section. EMA (cm2) was measured in the eye muscle cross-section. MS was scored on a scale of 1 thorough 9 (1 = trace, 9 = very abundant) according to the Korean Beef Marbling Standard.
Sequence analysis of bGH gene
Genomic DNA was extracted from white blood cells of 21 unrelated Hanwoo individuals using the phenolchloroform method (Sambrook et al., 2001). We sequenced 47 bp to 2,528 bp of the bGH gene (GenBank accession number, M57764) and the flanking regions to evaluate SNP variants using the BigDye Terminator (ver. 3.1) cycle sequencing kit (Applied Biosystems, Foster City, CA) with an ABI 3730XL DNA analyzer (Applied Biosystems). Six primer sets for amplification and sequencing analysis were designed based on the GenBank sequence using Primer3 software. Primer sequences are shown in Table 1. Sequence editing was performed by visual confirmation using Sequencher 4.6 software (Gene Codes Corp., Ann Arbor, MI).

Sequences of primers for sequencing the bovine growth hormone (bGH) gene in Hanwoo
Genotyping by single-base extension (SBE)
A primer set (GH-P1-F and GH1-P1-R) was designed to generate a 639-bp product that included four SNPs in the bGH gene promoter. Another primer set (GH-E1-F and GH-E1-R) was designed to generate a 328-bp product that included 679-bp SNP in the exon 1 region. A third primer set (GH-E2-F and GH-E2-R) was designed to produce a 536-bp amplicon that included 1,692-bp SNP in intron 3, while a primer set (GH-E3-F and GH-E3-R) was designed to obtain a 428-bp product that included four SNPs in exon 4 (Table 1). Primer extension was performed using a SNaPshot ddNTP Primer Extension Kit (Applied Biosystems, Foster City, CA). To purify the primer extension products, exonuclease 1 and shrimp alkaline phosphatase were added to the reaction mixtures. The samples were incubated at 37°C for 1 h and the reactions were stopped by incubating at 72°C for 15 min. The products were mixed with a Genescan 120 LIZ standard and HiDi formamide (Applied Biosystems) before being denatured at 95°C for 5 min. Electrophoresis was performed using an ABI PRISM 3130XL Genetic Analyzer and the results were analyzed using GeneMapper v.4.0 software (Applied Biosystems).
Statistical analysis
Heterozygosity, minor allele frequency (MAF), and Hardy-Weinberg equilibrium (HWE) were assessed using Haploviewer v4.2 (Barret et al., 2005). HWE was tested by comparing the expected and observed genotype frequencies using a chi-square test. Associations of growth and carcass traits for each SNP were analyzed with a mixed analysis of covariance (ANCOVA) linear model using SAS v9.1 (SAS Institute, Cary, NC). For growth traits, two fixed effect were fitted, year-season-birth place and SNP genotype. For carcass quality traits, an additional effect was included in the model, a covariate for age in days at the time of slaughter.
RESULTS AND DISCUSSION
The DNA segment of 2,482 bps that was located at 47 to 2,528 bp of bGH gene was sequenced, and a total of 10 SNPs were identified (Figure 1 and Table 2). Among the ten SNPs, four SNPs were located in the promoter region, i.e. 253 C>T, 303 C>T, 502 C>T, and 559 G>A. Results of association tests between the four SNPs and growth and carcass traits showed that 303 C>T and 559 G>A SNPs had significantly affected WT6 and eye muscle area (EMA), respectively at p = 0.05 level (Tables 3 and 4). The significant SNPs in the bGH gene promoter region may have limited efficiency as molecular markers, partly because the SNPs are located in the SINE/BovA2 repeat element, in which repetitive mutations occur frequently (Vaccarelli et al., 2008), suggesting that the SNPs does not strongly influence gene expression. Alternatively, the SNPs may affect growth and carcass traits due to great linkage disequilibrium with the causal variants that were closely located to the bGH SNPs (Ge et al., 2013). Kim et al. (2004) reported that a SNP at a promoter position -120 spanning a DraI restriction site was associated with 3-month weight and carcass weight in a Hanwoo population. However, our study did not confirm the SNP, partly because the SNP in Kim et al. (2004) was not detected with strong statistical evidence, i.e. comparison-wise p values were 0.025 and 0.041 for 3-month weight and carcass weight, respectively, which may be not confirmed in another random Hanwoo sample. Also, Kim et al. (2004) analyzed the association tests with estimated breeding values of the growth and carcass quality traits, while raw phenotypes were used in this study.
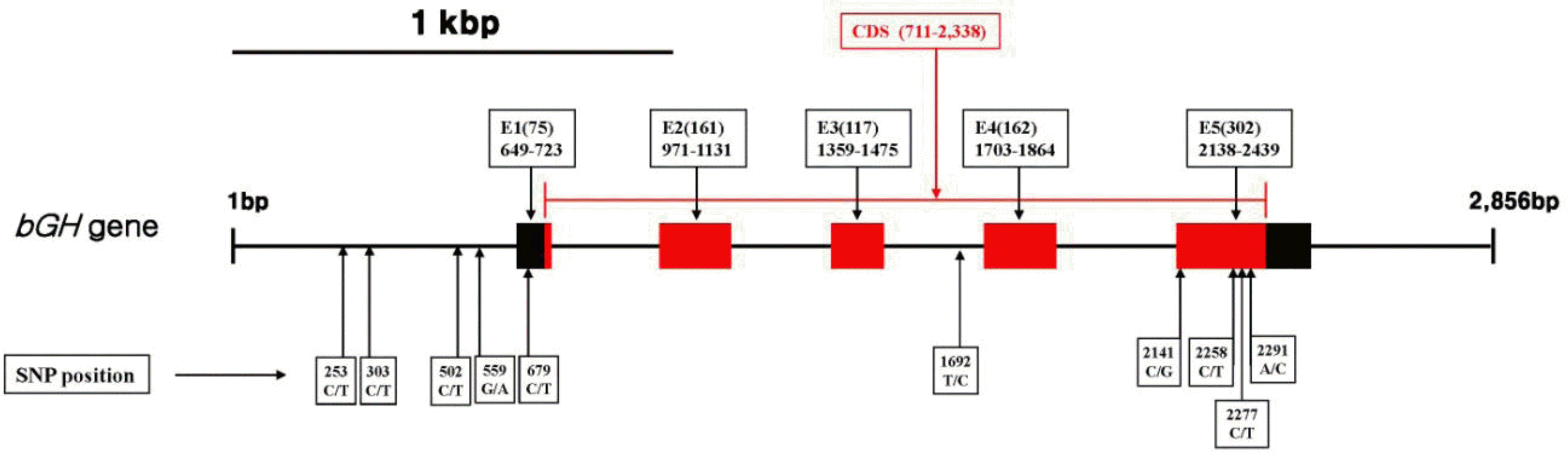
Position of SNPs in the bovine growth hormone (bGH) gene in Hanwoo.

Genotype and allele frequencies of SNPs within the bGH gene of Hanwoo

Least square mean and standard error of genotype effects of bGH gene for growth traits in Hanwoo

Least square mean and standard error of genotype effects of bGH gene for carcass quality traits in Hanwoo
In this study, 679 C>T SNP was found in exon 1, 1692 T>C SNP in intron 3, and four SNPs (2141 C>G, 2258 C>T, 2277 C>T, and 2291 A>C) in exon 5 of the bGH gene. Among the SNPs in the CDS region, the 2141 C>G, the 2258 C>T, and the 2277 C>T SNPs were non-synonymous causing amino acid substitution, while the 2291 A>C SNP was a silent mutation. Among the SNPs in exon 1 and intron 3, 679 C>T and 1692 T>C did not significantly influence any growth or carcass trait (Tables 3 and 4). These results are in accordance with the report of Yao et al. (1996), in which there was no significant association of the SNPs with milk yield, fat and protein content in Holstein bulls.
The 2141 C>G SNP was non-synonymous and induced a mutation from leucine (CTG) to valine (GTG), which significantly affected ADG at p = 0.05 level (Table 3), even if HWE for the SNP was significantly deviated from expectation (Table 2). The 2258 C>T SNP was non-synonymous, causing mutation of arginine to tryptophan in the process of transition from C to T. The SNP significantly affected ADG and CWT at p = 0.05 level (Tables 3 and 4). The genotype effect of the 2258 SNP on ADG was 0.74±0.01 for CC and 0.71±0.01 for CT, respectively (Table 3). For CWT, the estimates of CC and CT genotypes were 310.4±2.4 and 298.5±4.9, respectively (Table 4). For the SNP, Yoon et al. (2003) reported that MAF, Msp I (-), was low (0.00 to 0.054) in European Bos taurus species (Hereford, Angus, Charolais, Holstein, brown Swiss, Limousine, and Simmental), 0.043 to 0.229 in Asian Bos taurus breeds, except for Japanese black cattle (0.00), and 0.162 in Hanwoo. The MAF (C allele) of the 2258 SNP was 0.084 in this study (Table 4), which was lower than the frequency of the SNP in Yoon et al. (2003). This may be partly due to sampling effect, i.e. a small sample size (N = 242) in this study. For the 2258 C>T SNP, the high allele frequency of the favorable allele (C) in both European and Asian Bos taurus breeds indicate that selection for genetic improvement of ADG and CWT has been processed for the SNP or near the chromosomal region of the SNP.
There are some limitations in this association study. First of all, the sample size was small (N = 242), such that some significant SNPs for growth and carcass quality traits may not have been detected. Second, there may have been a chance of a false positive SNP, i.e. spurious SNPs with significant evidence that have no true effects on the tested traits. Some significant SNPs, e.g. the 303 C>T SNP for WT6 or the 2258 C>T SNP for CWT (Tables 3 and 4), had MAF less than 0.05, for which efficiency of marker-assisted selection would not be high.
Our results indicate that four SNPs in the bGH gene were associated with growth and carcass quality traits in Hanwoo (Tables 3 and 4). However, further study is needed to validate effects of the significant SNPs, before considering implementation of marker-assisted selection in Hanwoo commercial populations.
Acknowledgements
This research was supported by a grant titled as “Development of production technologies for high quality & nutrional values of beef in Hanwoo from the Technology Development Program for Agriculture and Forestry (Project No. 311016-3), Ministry of Agriculture, Forestry, and Fisheries, Republic of Korea. Jong-Joo Kim’s work was also supported by the Yeungnam University Research Grant 2012.