Essential oil mixture on rumen fermentation and microbial community – an in vitro study
Article information

Abstract
Objective
The objective of this study was to investigate the effects of essential oil mixture (EOM) supplementation on rumen fermentation characteristics and microbial changes in an in vitro.
Methods
Three experimental treatments were used: control (CON, no additive), EOM 0.1 (supplementation of 1 g EOM/kg of substrate), and EOM 0.2 (supplementation of 2 g EOM/kg of substrate). An in vitro fermentation experiment was carried out using strained rumen fluid for 12 and 24 h incubation periods. At each time point, in vitro dry matter digestibility (IVDMD), neutral detergent fiber digestibility (IVNDFD), pH, ammonia nitrogen (NH3-N), and volatile fatty acid (VFA) concentrations, and relative microbial diversity were estimated.
Results
After 24 h incubation, treatments involving EOM supplementation led to significantly higher IVDMD (treatments and quadratic effect; p = 0.019 and 0.008) and IVNDFD (linear effect; p = 0.068) than did the CON treatment. The EOM 0.2 supplementation group had the highest NH3-N concentration (treatments; p = 0.032). Both EOM supplementations did not affect total VFA concentration and the proportion of individual VFAs; however, total VFA tended to increase in EOM supplementation groups, after 12 h incubation (linear; p = 0.071). Relative protozoa abundance significantly increased following EOM supplementation (treatments, p<0.001). Selenomonas ruminantium and Ruminococcus albus (treatments; p<0.001 and p = 0.005), abundance was higher in the EOM 0.1 treatment group than in CON. The abundance of Butyrivibrio fibrisolvens, fungi and Ruminococcus flavefaciens (treatments; p< 0.001, p<0.001, and p = 0.005) was higher following EOM 0.2 treatment.
Conclusion
The addition of newly developed EOM increased IVDMD, IVNDFD, and tended to increase total VFA indicating that it may be used as a feed additive to improve rumen fermentation by modulating rumen microbial communities. Further studies would be required to investigate the detailed metabolic mechanism underlying the effects of EOM supplementation.
INTRODUCTION
Numerous ruminant nutritionists have tried to develop natural alternatives for antibiotics since the prohibition of their use in livestock industry in the European Union owing to the risk of antibiotic residues in animal products [1] and appearance of multi-drug resistant bacteria that might have a negative impact on human health [2]. Essential oils (EO) are secondary metabolites from plants that function in defense against herbivores and invading microbes and have been known to possess antimicrobial properties [3,4]. In addition, plant extracts are categorized in generally recognized as safe for human consumption; therefore, numerous studies have evaluated the efficacy of EO and essential oil compounds (EOC) with the aim of replacing antibiotic rumen modulators such as monensin, using an in vitro batch culture systems [2,5–7] and continuous culture systems [8–10].
In the previous studies, the separate use of individual EO (eugenol, thymol, and cinnamaldehyde) in an in vitro system has shown various effects, but results were inconsistent with respect to the modulation of ruminal fermentation [2,8,9,11]. Therefore, the use of essential oil mixtures (EOM) containing various EO has been widely assessed for the modulation of rumen fermentation, and the type and quantity of the EO in an EOM are the most important factors that determine their capability for modulating rumen fermentation [5]. Novatan (Techna, Couëron, France), a commercial EOM, has been developed for reducing excessive protein degradation in the rumen through the synergetic activity of EO (eugenol, thymol, and cinnamaldehyde) and microminerals (Na, Zn, and Mn) [12].
The aim of this study was thus to investigate the effects of dietary supplementation of a newly developed commercial EOM containing eugenol, thymol, and cinnamaldehyde, on rumen fermentation characteristics and changes in microbial diversity using an in vitro batch culture system.
MATERIALS AND METHODS
Animal use and the protocols for this experiment were reviewed and approved by the Animal Research Ethics Committee of Pusan National University.
Preparation of an experimental substrate
A commercial concentrate mix for fattening beef was purchased from a feed company (Easybio, Inc., Seoul, Korea). The detailed chemical composition was as follows: dry matter (DM), 900 g/kg; crude protein, 255 g/kg; neutral detergent fiber (NDF), 246 g/kg; acid detergent fiber (ADF), 116 g/kg; lignin, 35.3 g/kg; ether extracts, 48.1 g/kg; ash, 87.3 g/kg; Ca, 4.3 g/kg; P, 7.8 g/kg. The experimental substrates were dried at 60°C for 96 h, and ground in a cyclone mill (Foss Tecator Cyclotec 1093, Foss, Hillerød, Denmark) fitted with a 1 mm screen, for later use. A commercial EOM (NOVATAN, Techna, France) was used in this study as a feed additive. According to the manufacturer, the main constituents in the EOM were cinnamaldehyde, thymol, and eugenol.
Experimental treatments
A complete randomized block design was used for the experiment, with treatment as the main effect. Three experimental treatments were used, i.e., control (CON, no additive), EOM 0.1 (1 g EOM/kg of experimental substrate), EOM 0.2 (2 g EOM/kg of experimental substrate). The amount of EOM added was determined based on the manufacturer’s recommendation.
In vitro incubation
In vitro batch culture fermentation was carried out using rumen fluid, which was collected before the morning feeding from two cannulated Korean native cattle heifers (body weight 450±30 kg) at Center for Animal Science Research, Gyeongsang National University, Korea. Animals were fed a diet consisting of 600 g/kg timothy hay and 400 g/kg of a commercial concentrate mix. The rumen contents were mixed, transferred into a thermos bottle, and immediately transported to the laboratory. Rumen contents were strained through 4 layers of cheesecloth and mixed with 3× volumes of in vitro rumen buffer solution [13] under strictly anaerobic conditions. Approximately 0.9 g DM of the ground experimental substrate and a corresponding quantity of EOM for each treatment were moved into pre-weighed filter bags (F57, Ankom Technology, Macedon, NY, USA). All bags were heat-sealed and transferred into empty 125 mL serum bottles, to which 50 mL of rumen fluid and buffer mixture were transferred accompanied by continuously flushing with O2-free CO2 gas. The bottles were sealed with butyl rubber stoppers and aluminum caps and incubated on a rotary shaker (JSSI-300T, JS Research Inc., Gongju, Korea) at 125 rpm, for 12 and 24 h at 39°C. Three replicates were prepared for each treatment and each incubation time. After each incubation period, the three replicates of each treatment were removed from the incubator, and total gas production was measured using a pressure transducer (Sun Bee Instrument Inc., Seoul, Korea) as described by Theodorou et al [14]. Subsequently, the bottles were immediately placed on ice and the caps were opened to stop the fermentation; the filter bags were removed from bottles and rinsed with flowing water until the water ran clear. The pH of the culture fluid was measured using a pH meter (FP20, Mettler Toledo, Columbus, OH, USA). Sample fluid (5 mL) was stored at −80°C until rumen microbial population analysis. The remaining culture fluid was then centrifuged at 15,000×g for 10 min at 4°C and stored at −20°C, until the analysis of volatile fatty acids (VFA) and ammonia nitrogen (NH3-N) concentrations. The washed bags were dried at 55°C for 48 h and weighed to measure in vitro DM degradability (IVDMD). The NDF content was assessed for the weighed bags using a modified version of the micro-NDF method proposed by Pell and Schofield [15] to evaluate in vitro NDF degradability (IVNDFD).
Total DNA extraction
Collected rumen fluid samples were thawed in a refrigerator at 4°C, and centrifuged at 16,000×g and 4°C for 5 min. The supernatant was discarded, and the remainder was resuspended in 800 μL of 1× phosphate-buffered saline solution to wash the pellet. This was recentrifuged (16,000×g, 4°C, 5 min) and the supernatant was discarded. Total DNA was extracted from the washed pellet using a NucleoSpin DNA Stool kit (MACHEREY-NAGEL GmbH Co. KG, Düren, Germany), according to the manufacturer’s instructions. The concentration and purity of total DNA were measured using a NanoDrop (ND-1000, Thermo Fisher, Waltham, MA, USA).
Conventional polymerase chain reaction for validation of primer and real-time polymerase chain reaction
The information on primer sequences of rumen microbes was collected from previous studies [16,17] and presented in Table 1. Conventional polymerase chain reaction (PCR) was performed to validate the specificity of the primer in 50 μL reaction volumes using AccuPower PCR PreMix K-2013 (Bioneer, Daejeon, Korea), by adding 2 μL each of forward and reverse primers, 2 μL of genomic DNA and 44 μL of PCR grade water. Reactions were performed using a C1000 Thermal cycler (Bio-Rad, Hercules, CA, USA) under the following conditions: initialization for one cycle at 95°C for 10 min; 35 cycles each for denaturation at 95°C for 30 s, annealing at 60°C for 30 s, and elongation at 72°C for 30 s; and final elongation at 72°C for 5 min. The PCR products were analyzed by electrophoresis using 1.5% agarose gel impregnated with Staining STAR (DYNEBIO INC, Seongnam, Korea), to confirm a single specific band and the absence of primer dimer products.

Polymerase chain reaction primers used in this study
Real-time quantitative PCR assays were performed on a CFX 96 Touch system (Bio-Rad Laboratories, lnc. USA). Reactions were performed in triplicate with reaction volumes of 20 μL using optical reaction plates sealed with optical adhesive film. Each reaction mixture contained 0.5 μL 10 mM dNTP Mix (BioFACT, Daejeon, Korea), 2 μL 10× buffer (BioFACT, Korea), 1 μL 40 ng genomic DNA, 1 μL forward primer (10 pmol), 1 μL reverse primer (10 pmol), 0.1 μL Taq polymerase (BioFACT, Korea), 1 μL Evagreen (SolGent Co., Ltd., Daejeon, Korea) and 13.4 μL PCR grade water. Real-time PCR was carried out according to the manufacturer’s instructions, as follows: initialization for one cycle at 95°C for 10 min; 40 cycles each for denaturation at 95°C for 30 s, annealing at 60°C for 30 s and elongation at 72°C for 30 s; and final elongation at 72°C for 5 min. Fluorescence was recorded at the end of each denaturation and extension step, and specificity of amplicon was confirmed via dissociation curve analysis of PCR end products by increasing the temperature at a rate of 1°C after every 30 s, from 60°C to 95°C. The threshold cycle (Ct) was analyzed and transformed using a standard curve with CFX manager software (Bio-Rad, USA).
Relative genomic DNA abundance of genes
To evaluate the efficiency (E) of amplification of each primer set, a 6-point standard curve and a nontemplate control (NTC) were run for each sample to test the relative expression levels. The standard curve was generated using genomic DNA (40 μg/reaction mixture), which was serially diluted. Amplification efficiency was calculated from the slope of the standard curve generated by plotting the threshold cycle (Ct) against logarithmic values of different DNA concentrations using the equation, E = 10−1/slope. The gene ‘general bacteria’ was used as a reference gene. Relative quantification of microbes was carried out using the 2−ΔΔCt method as follows [18,19]:
Statistical analysis
Statistical analysis of the data was performed using the MIXED procedure of SAS 9.3 (SAS institute Inc., NC, USA). The fixed effect in this model was a feed treatment. Orthogonal contrast was used to analyze linear and quadratic effect when the level of EOM supplementation increased. Differences among treatments were compared using Tukey’s range test if a significant effect was observed. Statistical significance was declared at p<0.05 and a trend was speculated at 0.05≤p<0.10.
RESULTS
The effects of EOM on rumen fermentation characteristics are presented in Table 2. After incubating for 12 h, there was no significant difference in IVDMD, IVNDFD, pH, and NH3-N concentration, among treatments. After incubating for 24 h, The IVDMD was significantly changed in a quadratic manner (p = 0.008), and the IVNDFD tended to increase linearly when the EOM was supplemented (p = 0.068). The pH was quadratically changed in when EOM was increasingly added (p = 0.032). No significant effect on NH3-N concentrations was detected at 12 h incubation but, the highest NH3-N concentration was observed in the group of EOM 0.2 at 24 h fermentation (p = 0.032 for Trt, and p = 0.013 for linear).
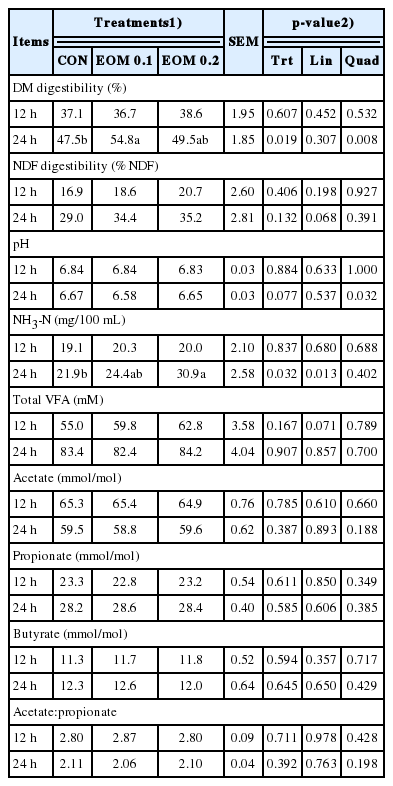
Fermentation characteristics after in vitro incubation of treatments using strained rumen fluid
Dietary addition of EOM tended to increase the total VFA concentration linearly after 12 h incubation (p = 0.071). There was no significant difference in total VFA concentration among treatments after 24 h incubation. Significant changes in the proportion of each VFA, and acetate to propionate ratio was not observed in any incubation times.
The EOM 0.1 treatment changed the fold change of Selenomonas ruminantium (p<0.001) Fibrobacter succinogenes (p = 0.047), and Ruminococcus albus (p = 0.005) in a quadratic manner (Table 3). EOM 0.2 treatment increased fold change of ciliate protozoa (p<0.001), Butyrivibrio fibrisolvens (treatment, linear, and quadratic effect: p<0.001, 0.002, and 0.007, respectively), fungi (treatment and linear effect: p = 0.001 and <0.001, respectively), and Ruminococcus flavefaciens (treatment, linear, and quadratic effect: p<0.001, <0.0001, and, 0.002, respectively). Prevotella ruminicola abundance tended to increase in a linear manner by EOM supplementation (p = 0.080).
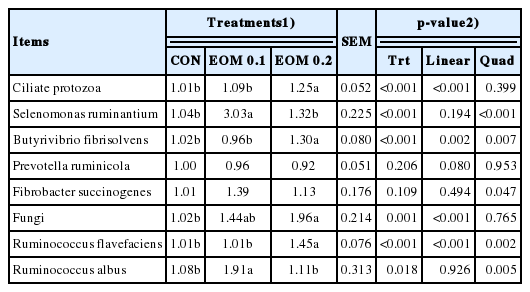
Fold changes of rumen microbial populations after 24-h in vitro incubation of treatments using strained rumen fluid
DISCUSSION
The EO supplementation in ruminant diets can change rumen microbial activity and fermentation characteristics because of their antimicrobial activity. Dorman and Deans [3] also reported that the inconsistent rumen fermentation results following the dietary addition of EO might be attributed to the type and structural configuration of EO when different kinds of EO are administrated together. Therefore, numerous researchers have been developing new EOM candidates for useful modulation of rumen environment. In this study, newly developed EOM containing eugenol, thymol, and cinnamaldehyde was evaluated for efficacy to improve rumen fermentation pattern by in vitro batch culture.
Results from this study revealed that appropriate dosage of EOM improves rumen degradability and microbial activity. Cinnamaldehyde consisting EOM has been known to bind to feed proteins and inhibit the amino acid decarboxylase in Enterobacter aerogenes by disturbing energy metabolism of bacterial cells [21]. However, NH3-N significantly increased in response to supplementation of the high dose of EOM. Ammonia nitrogen is an end product generated by microbial fermentation of protein sources in the feed. In the previous studies, the NH3-N was not affected by the addition of BEO [9,22] and, Benchaar et al [2] did not observe any significant changes in NH3-N concentrations when EOs or EOCs were added to substrates. Furthermore, Busquet et al [6] reported that the addition of eugenol and cinnamaldehyde to substrates at over doses of 300 mg/L, decreased NH3-N concentrations. Considering that our dosage level of EOM 0.2 was 30 mg/L, it is postulated that the dosage level of EOM in this study might be too low to modulate microbial degradative action on protein sources. Newbold et al [23] reported that defaunation of protozoa significantly decreased the rumen NH3-N concentration (−26%, p<0.001), indicating that, in this study, the increase of relative fold change of ciliate protozoa with an increase in the level of EOM supplementation might be linked with a higher concentration of NH3-N in EOM treatments than those in CON.
In present study, an increased IVDMD and IVNDFD were observed in groups treated with EOM after 24-h incubation (Table 3). Regarding with IVDMD, Benchaar et al [2] observed a significant decrease, when diets were supplemented with carvacrol (400 mg/L) and eugenol (800 mg/L) (p<0.05), but other EO (cinnamon leaf oil, clove leaf oil, sweet orange oil, oregano oil, and thyme oil) and EOC (cinnamaldehyde and thymol) had no effect on IVDMD in an in vitro batch culture system after 24 h incubation. Castillejos et al [9] reported that IVDMD was not changed by supplementation of EOM containing thymol, limonene, and guaiacol in an in vitro continuous culture system. Benchaar et al [2], Castillejos et al [7], and Fraser et al [10] observed a decrease in IVNDFD, and Benchaar et al [2] explained that fibrolytic bacteria might be sensitive to high concentrations of phenolic compounds in EO thereby fiber digestibility might be impaired by EO supplementation. Fibrobacter succinogenes, Ruminococcus flavefaciens, and Ruminococcus albus are the bacteria constituting 33.0%, 2.6%, and 46.0%, respectively, of the rumen fibrolytic bacteria [24,25]. Rumen ciliate protozoa have been known to have fibrolytic activity [23], and the fungi in the rumen have also been considered to produce fibrolytic enzymes [26]. In the present study, EOM 0.1 significantly increased the fold change of Ruminococcus albus (p = 0.005) and increased that of Fibrobacter succinogenes in a quadratic manner (p = 0.047). EOM 0.2 significantly increased the fold change of ciliate protozoa (p<0.001), Butyrivibrio fibrisolvens (p<0.001), fungi (p = 0.001), and Ruminococcus flavefaciens (p<0.001). Thus, the EOM used in this study may promote fibrolytic bacterial, protozoa, and fungal growth by unknown synergistic effect between chemical compounds and the main EO in the newly developed EOM, thereby improving ruminal degradability.
Regardless with increased digestibility, total VFA and those proportions after 24 h incubation were not changed by EOM supplementation. We expected that total VFA concentration would increase because the increase in nutrient digestibility and changes of central ruminal bacterial abundance were observed at 24 h incubation. Several studies reported that total VFA concentrations or their proportion were not affected by the supplementation of EO [26,27] irrespective of changes in ruminal nutrient digestibilities and the reason was not explained clearly as well. We postulated that the increase in protozoal abundance by EO addition might be linked with the increase in fiber digestibility without changes of VFA production because they engulf feed particles and ruminal bacteria thereby the bacterial fermentation contribution to VFA production would be impaired [22].
CONCLUSION
The addition of newly developed EOM to an in vitro batch culture affected ruminal fermentation characteristics and microbial fold changes. An increased IVDMD and IVNDFD were observed when EOM was added. It was suggested that the newly developed EOM may be used as a feed additive to improve rumen fermentation. Further studies are required to investigate the detailed metabolic mechanism underlying the effects of EOM supplementation.
ACKNOWLEDGMENTS
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. PJ01329201)” Rural Development Administration, Republic of Korea.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript. Kwon I is an employee of EASY BIO, Inc..