Effects of zinc bearing palygorskite supplementation on the growth performance, hepatic mineral content, and antioxidant status of broilers at early age
Article information
Abstract
Objective
This study was conducted to investigate effects of zinc (Zn) bearing palygorskite (ZnPal) supplementation on growth performance, hepatic mineral content, and antioxidant status of broilers at early age.
Methods
A total of 240 1-day-old Arbor Acres broiler chicks were allocated into 5 treatments with 6 replicates of 8 chicks each. Birds in 5 treatments were fed a basal diet supplemented with 0 (Control group; Analyzed Zn content: 81 mg/kg), 20, 40, 60, and 80 mg/kg Zn as ZnPal for 21 days, respectively. Blood, liver and intestinal mucosa were collected at 21 days of age.
Results
Treatments did not affect growth performance of broilers during the 21-day study (p> 0.05). The contents of hepatic Zn and magnesium (Mg) were linearly increased (p<0.001) by ZnPal supplementation. ZnPal inclusion linearly (p = 0.007) reduced malondialdehyde (MDA) concentration in serum. The activity of total superoxide dismutase (T-SOD) in liver increased linearly (p = 0.001) with concentration of ZnPal in diet. ZnPal inclusion linearly (p = 0.036) and quadratically (p = 0.005) increased T-SOD activity, and linearly (p = 0.012) increased copper/zinc superoxide dismutase (Cu/Zn SOD) activity in jejunal mucosa. The maximum responses of hepatic and jejunal antioxidant enzymes activities (T-SOD and Cu/Zn SOD) were found when supplementing the basal diet with 60 mg/kg Zn as ZnPal. Furthermore, ZnPal supplementation quadratically (p = 0.001) increased Cu/Zn SOD activity in ileal mucosa, and its maximum activity was observed in the diet supplemented with 20 mg/kg Zn as ZnPal.
Conclusion
ZnPal supplementation did not alter growth performance of broilers. Dietary ZnPal inclusion could increase concentrations of hepatic trace minerals (Zn and Mg) and inhibit lipid peroxidation by reducing serum MDA accumulation, with the optimal dosage of Zn from ZnPal being 80 mg/kg diet (analyzed Zn content in the diet: 165 mg/kg), and 60 mg/kg Zn as ZnPal (analyzed Zn content in the diet: 148 mg/kg) was the optimum dosage for broilers to achieve maximum antioxidant enzyme activities.
INTRODUCTION
The reactive free radicals, like reactive oxygen species (ROS), are routinely produced during metabolism in animals. Generally, the amounts of these radicals maintain a balanced status and are not harmful for metabolism and health of animals in the presence of an effective antioxidant defense system [1]. However, when the over production of ROS exceeds the defence capacity of the host’s antioxidant system, it will trigger lipid peroxidation that would induce oxidative damages to DNA and proteins, disintegrate the cellular membrane, disturb the balance between oxidants and antioxidants, and eventually induce the occurrence of oxidative stress, which is often regarded as one of the causes of several pathologies that affect poultry growth [1,2]. Antioxidant defense system is comprised of non-enzymatic and enzymatic antioxidant system. The enzymatic antioxidant system includes glutathione peroxidase, catalase, copper/zinc superoxide dismutase (Cu/Zn SOD), and small molecular antioxidants as reduced glutathione, and this antioxidant system can efficiently protect the integrity of cellular membrane and maintain normal function of tissues through removing the excessive oxygen-derived free radicals and oxidizing products [2].
Zinc (Zn), as an essential trace element, acts both structurally and catalytically in various metalloenzymes, and therefore is indispensable for the growth, development, reproduction, and metabolic activities of both avian and mammalian species [3–5]. For broilers, Zn is usually supplemented to diets as Zn sulfate or Zn oxide to meet the requirement of birds since many natural feed ingredients are marginally deficient in Zn [6]. Zn is the cofactor of Cu/Zn SOD, a major antioxidant enzyme [7], and plays a key role in the antioxidant defense system [8,9]. Bun et al [5] reported that supplementation of organic Zn (from 20 to 60 mg/kg diet) to a corn–soybean basal diet (29.6 mg Zn/kg diet) ameliorated oxidative stress of broilers by increasing the activities of antioxidant enzymes including Cu/Zn SOD, and inhibiting lipid peroxidation, irrespective of whether birds were healthy or challenged with Eimeria tenella. Similarly, Zhao et al [10] observed that Zn oxide nanoparticles inclusion improved the antioxidant capacity of broilers, as demonstrated by increased Cu/Zn SOD activity and reduced malondialdehyde (MDA) accumulation in both serum and liver. Also, it has been demonstrated that Zn supplementation in the form of either inorganic or organic source improved the oxidative status in quail under heat stress [11,12].
Palygorskite (Pal) is a naturally available magnesium alumi num silicate clay mineral with adsorption property, adhesive ability and cation exchange capacity [13]. In animal nutrition, Pal, either as a feed ingredient or feed additive, has been widely incorporated in animal feed [14,15]. Due to the characteristics of Pal aforementioned, Zn bearing Pal (ZnPal) recently has been prepared using solid state ion exchange method [16]. In broilers, previous studies have shown that ZnPal could be used as a potential Zn source with a higher bioavailability than that of Zn sulfate [16]. In our previous study, the supplementation of ZnPal improved meat quality and its mineral composition, and enhanced muscle antioxidant capacity, whereas it did not affect the growth performance of broilers at 42 days of age [17]. In an in vivo study, Tang et al [18] found that Zn bearing clinoptilolite improved anti-oxidative status in the intestine of broilers as evidenced by increased Cu/Zn SOD activity and reduced accumulation of MDA, the end product of lipid peroxidation. Considering the similar physical and chemical characteristics among clays and the results of our previous study [17], we then hypothesized that the inclusion of ZnPal to diet would also improve the antioxidant capacity in the intestine and liver of broilers, and therefore the current study was conducted to investigate the effects of ZnPal supplementation on the growth performance, hepatic mineral content and antioxidant status in the liver and intestine of broilers at early age.
MATERIALS AND METHODS
Preparation of ZnPal
The Pal was kindly offered by Jiangsu Sinitic Biotech Co., Ltd. (Xuyi, Jiangsu, China) and passed through a 200-mesh sieve (diameter, 0.074 mm). The main chemical composition of Pal determined by a Minipal 4X-ray fluorescence spectrometer (PAN analytical Co., Eindhoven, Netherlands) are listed in the following: SiO2, 59.11%; MgO, 12.75%; Al2O3, 10.31%; CaO, 7.42%; Fe2O3, 6.21%; Na2O, 1.27%; and K2O, 1.19%.
ZnPal was prepared according to method of solid state ion exchange as previously described by Yan et al [16]. Namely, Pal was firstly calcinated at around 300°C for 1 h in the muffle oven. After cooling down, Pal was mixed with ZnCl2 (ZnCl2 ≥98.0%; 4:1, wt/wt) purchased from Nanjing Chemical Reagent Co., Ltd. (Nanjing, Jiangsu, China) in a stainless steel blade grinder (HJ-6A, Changzhou Guohua Electric Appliance Co., Ltd., Changzhou, Jiangsu, China). The mixture was subsequently calcinated at 300°C for 3 h in a muffle oven. After cooling down to room temperature, the mixture was washed repeatedly by deionized water until there was no white deposition generated in the washed solution when swigged with 0.1 mol/L AgNO3 solution. Finally, the washed mixture were collected and dried at around 105°C for 2 h in an air oven, and then passed through a 200-mesh sieve after cooling down. The amount of Zn adsorbed by Pal was 47.15 mg/g.
Experimental design, diets and management
All procedures were approved by Nanjing Agricultural University Institutional Animal Care and Use Committee. Experimental design, diets and management in this study was the same to that of our previous study [17]. Briefly, a total of 240 1-day-old Arbor Acres broiler chicks (initial weight, 36.71±0.18 g) obtained from a commercial hatchery were randomly allocated into 5 dietary treatments with 6 replicates (cages) of 8 chicks each (4 males and 4 females/cage). Birds in the 5 treatments were given a basal diet supplemented with 0 (Control group), 20, 40, 60, and 80 mg Zn/kg diet as ZnPal for 21 days, respectively. The formulation and calculated nutrient level of basal diet are shown in Table 1. All birds were placed in wired cages and housed in an environmentally controlled room with continuing light. Temperature was maintained at 32°C to 34°C for the first 3 days and then reduced by 2°C to 3°C per week until a final temperature of 26°C was achieved. Birds were allowed ad libitum access to mash feed and water.
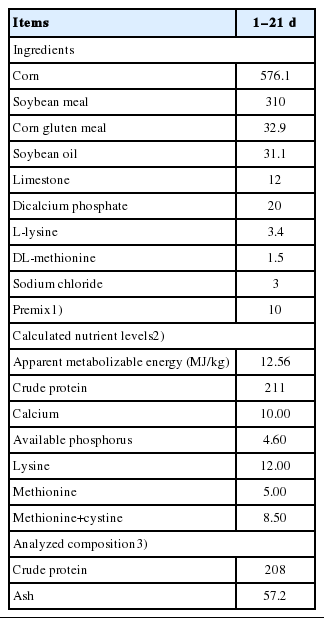
Composition and nutrient level of basal diet (g/kg, as fed basis unless otherwise stated)
Growth performance
At 21 days of age, birds were weighed by cage (replicate) after a 12-h fasting period to determine final body weight, and feed consumptionin each cage during the 21 days was recorded by subtracting residual feed from the total feed provided. Average daily feed intake (ADFI) was measured by dividing average feed consumption (feed consumption divided by bird number) by the days (21 day). Average daily gain (ADG) was calculated by dividing average body weight gain (average final body weight subtracted average initial body weight) by the days. During the experiment, dead birds were recorded and weighed. Feed/gain ratio (F:G) was calculated by dividing average feed intake by average weight gain, including weight gain of any dead birds.
Sample collection
At 21 days of age, 6 birds per treatment (1 broiler per replicate) were selected and weighed. Blood samples (around 8 mL) were taken from the wing vein before slaughter, and serum was separated by centrifugation at 3,000 g for 15 min at 4°C, which was then stored at −20°C until analysis. After that, birds were euthanized by cervical dislocation. After dissection, a portion of the right lobe of liver samples were excised, washed twice with phosphate buffer solution (pH = 7.4), dried with filter paper, minced, snapped frozen in liquid nitrogen, and then stored at −20°C for subsequent analysis. Meanwhile, jejunum (from the end of pancreatic loop to the Meckel’s diverticulum) and ileum (from Meckel’s diverticulum to the ileocecal junction) were removed of the mesentery and placed on a chilled stainless steel tray immediately, which were then opened longitudinally, and the intestinal digesta were flushed with ice-cold phosphate buffered saline. After that, the jejunal and ileal mucosa were scrapped carefully using a sterile glass microscope slide, rapidly frozen in liquid nitrogen and then stored at −80°C for further analysis.
Hepatic mineral content determination
The contents of Zn, Cu, manganese (Mn), iron (Fe), and magnesium (Mg) in the liver and feed samples were determined according to the method described by Tang et al [19], which was further optimized by Yan et al [16]. Briefly, 0.50±0.02 g feed or 2.0±0.1 g fresh liver samples were weighed into a glass digestion tube, mixed with 10 mL of a mixture acid of nitric acid and perchloric acid (4:1, vol/vol) at room temperature for 12 h, and then digested on a heating block (LabTech DigiBlock Digester, EHD36, Labtech Co., Ltd., Beijing, China) to acquire clear digested solution. The procedure of digestion was in the following: 90°C for 30 min; 120°C for 30 min; 160°C for 120 min; 180°C for 180 min. After that, digested solutions were diluted with ultra-pure water (high performance liquid chromatography-grade water) that was prepared using a Milli-Q system (Millipore, Molsheim, France) to a final volume of 25 mL. The final solutions were analyzed for mineral content by an inductively coupled plasma mass spectrometry (Optimal 2100 DV, Perkin-Elmer-Sciex, Norwalk, NY, USA). The operating conditions were: power, 1,300 W; plasma gas flow rate, 12 L/min; auxiliary gas flow rate, 0.2 L/min; nebuliser gas flow rate, 0.55 L/min; sample flow rate, 1.5 mL/min; sample uptake rate, 1.0 mL/min. Validation of the minerals analysis was conducted using a certified bovine liver powder (GBW (E) 080193; National Institute of Standards and Technology, Beijing, China) as a standard reference material. Analyzed mineral element content in the diets is presented in the Table 2.

Analyzed trace minerals content in the diets during 1 to 21 days (mg/kg)
Measurement of serum antioxidant parameters
Serum samples were analyzed for the activities of total superoxide dismutase (T-SOD, # A001-1) and Cu/ZnSOD (# A001-2), and MDA (# A003-1) content using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions. Briefly, the content of MDA was measured by barbiturate thiosulfate method, and the activity of SOD was determined by the hydroxylamine method. One unit of SOD was defined as the amount of SOD required to produce 50% inhibition of the rate of nitrite production at 37°C.
Measurement of hepatic and intestinal mucosal antioxidant parameters
Around 0.30±0.01 g of liver or intestinal mucosa samples were homogenised (1:9, wt/vol) with ice-cold 154 mmol/Lsterile sodium chloride solution using an Ultra-Turraxhomogeniser (Tekmar Co., Cincinatti, OH, USA) and then centrifuged at 4,000 g for 15 min at 4°C. The total protein concentrations were determined using a corresponding diagnostic kit (Nanjing Jian cheng Bioengineering Institute, China) according to the instructions of the manufacture. The determination methods of SOD activity and MDA concentration were the same as the measurements of serum samples. The concentration of MDA was expressed as nanomole per milligram protein of sample. All results were normalized to total protein concentration in each sample for inner-sample comparison (Value of enzyme activity [T-SOD and Cu/Zn SOD] or MDA concentration in each sample was divided by corresponding total protein concentration).
Statistical analysis
Data were analyzed by one-way analysis of variance using SPSS (2008) statistical software (Ver. 16.0 for Windows, SPSS Inc., Chicago, IL, USA). The differences among treatments were examined by Tukey’s test, which were considered to be significant at p<0.05. Polynomial contrasts were used to determine the linear and quadratic effects of dietary ZnPal inclusion level. Means and standard errors were presented.
RESULTS
Growth performance
Treatments (Table 3) did not alter growth performance of broilers (ADFI, ADG, and F:G) during the 21-day study (p>0.05).
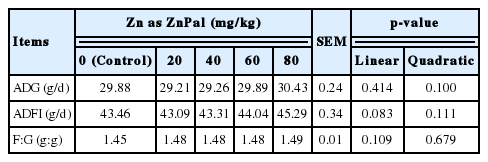
Effects of zinc bearing palygorskite supplementation on the growth performance of broilers from 1 to 21 days of age
Hepatic mineral content
As indicated in Table 4, the contents of Zn and Mg in the liver were linearly (p<0.001) increased by ZnPal addition in the diet, and the maximum concentrations of Zn and Mg in the liver were observed when supplementing the basal diet with 80 mg/kg Zn as ZnPal (p<0.05). However, no significant differences were observed in the contents of hepatic Fe, Cu, or Mn among treatments (p>0.05).
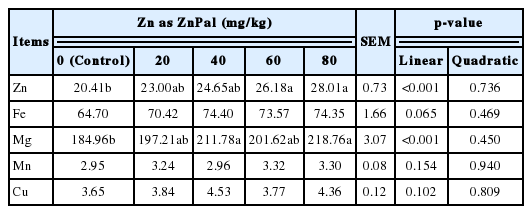
Effects of zinc bearing palygorskite supplementation on the hepatic minerals content of broilers at 21 days of age (mg/kg fresh sample)
Serum antioxidant status
ZnPal inclusion linearly (p = 0.007) reduced the MDA concentration in serum (Table 5) of broilers, with the lowest MDA content being found in the diet supplemented with 80 mg/kg Zn in the form of ZnPal (p<0.05). Birds exhibited similar T-SOD and Cu/Zn SOD activities in the serum among groups (p>0.05).
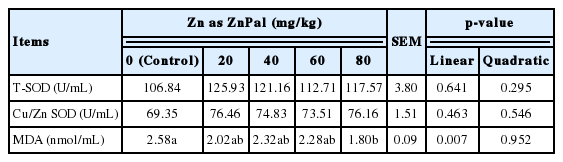
Effects of zinc bearing palygorskite supplementation on the serum antioxidant capacity of broilers at 21 days of age
Hepatic antioxidant status
ZnPal supplementation (Table 6) linearly (p = 0.001) increased the activity of T-SOD in the liver of broilers, and its maximum response was observed in the diet supplemented with 60 mg/kg Zn as ZnPal (p<0.05). However, there were no differences in the activity of Cu/ZnSOD or MDA concentration (p>0.05) among groups.
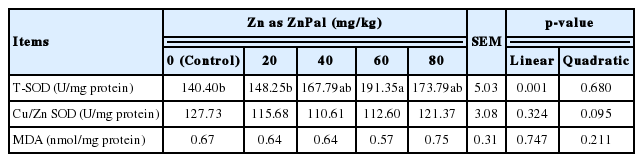
Effects of zinc bearing palygorskite supplementation on the hepatic antioxidant capacity of broilers at 21 days of age
Intestinal mucosal antioxidant status
Supplementation of ZnPal (Table 7) linearly (p = 0.036) and quadratically (p = 0.005) increased the activity of T-SOD, and linearly (p = 0.012) increased the activity of Cu/Zn SOD in jejunal mucosa. Broilers fed the diet supplemented with 60 mg/kg Zn in the form of ZnPal exhibited the highest activities of jejunal T-SOD and Cu/Zn SOD (p<0.05). Similarly, ZnPal supplementation quadratically (p = 0.001) increased the activity of Cu/ZnSOD in ileal mucosa, and the maximum activity of which was found in the diet supplemented with 20 mg/kg Zn (p<0.05). No significant differences were observed in the activity of ileal T-SOD or intestinal mucosal MDA content (p>0.05).
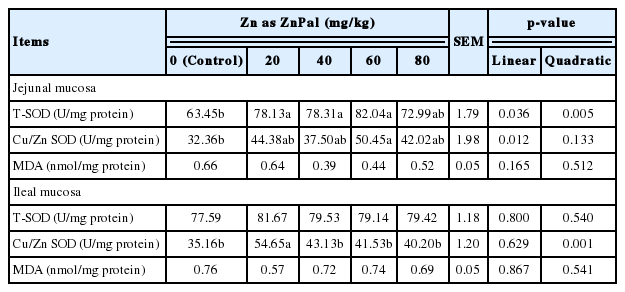
Effects of zinc bearing palygorskite supplementation on the mucosal antioxidant capacity of broilers at 21 days of age
DISCUSSION
In broilers, Bun et al [5] showed that the incorporation of an organic Zn source (methionine hydroxyl analog–Zn chelate) to a corn–soybean basal diet (29.6 mg Zn/kg diet) from 0 to 60 mg/kg did not alter weight gain, feed intake and feed/gain ratio of healthy birds or birds challenged with Eimeria tenella. Similarly, in our present study, ZnPal supplementation did not affect the growth performance (ADG, ADFI, and F:G) in broilers from 1 to 21 days of age, and it was partially consistent with the results of Yan et al [16] who reported that broilers fed a basal diet (35.2 mg Zn/kg diet) supplemented with 0, 20, 40, 60, or 80 mg/kg Zn in the form of ZnPal exhibited similar growth performance (weight gain, feed intake, and feed conversion ratio) at 21 days of age. Actually, the similar growth performance of broilers among groups in this study lasted until the end of study (42 day) as reported in our previous study [17]. For broilers, Huang et al [20] have shown that the optimum concentration of Zn in chicken diet from hatch to 21 days of age is 84 mg/kg. Additionally, Sunder et al [21] demonstrated that even 29 mg/kg Zn present in corn-soybean meal basal diet was capable of supporting performance in broilers up to 4-wk of age. In the current study, the analyzed content of Zn in the basal diet was 81 mg/kg (Table 2), and thus the similar growth performance among treatments in our present study may be due to the Zn level in the basal diet, which was able to meet the growth requirements of chicks at early age. In contrast, Liu et al [22] reported that higher feed intake and weight gain were observed in broilers fed diet supplemented with 60, 120, or 180 mg/kg Zn in the forms of ZnSO4, Zn amino acid and Zn proteinate when compared with those fed the basal diet (35.6 mg/kg) devoid of supplemental Zn, and authors concluded that 90 mg/kg Zn was sufficient to reach an optimal growth performance in broilers since growth performance of broilers were similar among the Zn-supplemented groups. Thus, this discrepancy among studies was most likely to result from level of Zn in the basal diet (35.6 mg/kg) of the study of Liu et al [22], unlike the present study (81 mg/kg), which could not meet birds’ requirement for growth performance.
In poultry nutrition, Tang et al [19] have demonstrated that Zn concentration in the liver, pancreas and tibia in broilers at 14 days of age exhibited a linear response to an increasing concentration of Zn bearing zeolite clinoptilolite, an alternative Zn source to Zn sulfate. In this study, the supplementation of ZnPal also linearly increased Zn accumulation in the liver of broilers, indicating that Zn adsorbed by Pal was ready for use by broilers, which in turn enable ZnPal to be a potential Zn source. Similarly, Yan et al [16] have shown that dietary ZnPal or Zn sulfate supplementation increased Zn retention in the pancreas and tibia of broilers at both 21 and 42 days of age. The increased hepatic Zn concentration resulting from ZnPal supplementation observed in this study may be associated with its modulation on the mRNA abundances of metallothioneins and Zn transporters, the key mediators for Zn adsorption and transportation, as previously reported in broiler chickens [19] and laying hens [23] when supplemented with Zn bearing zeolite clinoptilolite. Dietary ZnPal supplementation linearly increased hepatic Mg in this study, and it was consistent with our previous results in the breast muscles of broilers [17]. The beneficial effects of Zn bearing zeolite or montmorillonite on the intestinal morphology of animals (one of the key factors for nutrient adsorption) have been observed previously [18,24]. In the current study, the analyzed Mg in the diets was similar among the five groups as indicated in the Table 2, and therefore the enhanced Mg accumulation in the liver may suggest a promoted Mg bioavailability in the ZnPal. Considering the similar chemical and physical characteristics among clays, the enhanced Mg accumulation in the liver may be in related to the protective effect of ZnPal in the intestinal integrity.
The SOD, one of the most important antioxidant enzymes in the antioxidant defense system, ubiquitously exists innumerous cells of animals that could catalyze the disproportionation of superoxide to molecular oxygen and peroxide, and therefore is critical for protecting the cell against the toxic products of aerobic respiration [7,25]. As one of the isoforms of SOD, the enzyme Cu/Zn SOD has two identical subunits, and each subunit contains a metal cluster, the active site, constituted by a Cu and a Zn atom bridged by a histamine residue [26]. Cu/Zn SOD enzyme can coordinate the removal of oxygen free radicals and is believed to play a major role in the first line of antioxidant defense [7,26]. MDA is one of the end-products of lipid peroxidation by ROS, and the level of which is often regarded as an indicator of lipid peroxidation [27]. In this study, dietary Zn supplementation as ZnPal inhibited the generation of MDA in the serum, and increased the activities of T-SOD and Cu/Zn SOD in the tissues (liver and intestine), indicating that the inclusion of ZnPal improved the antioxidant capacity of broilers at early age. Likewise, in an in vivo study, Bun et al [5] have shown that the inclusion of organic Zn improved oxidative status of broilers (irrespective of their health status) by increasing the activities of Cu/Zn SOD and glutathione peroxidase, and reducing MDA accumulation in the serum. Moreover, Zhao et al [10] also found that the incorporation of Zn oxide nanoparticles enhanced the antioxidant capacity of broilers. In this study, the improved oxidative status was in agreement with the simultaneously increased Zn retention in the liver, which in turn suggested that the enhanced antioxidant capacity may result from elevated Zn accumulation. The mode action of Zn in antioxidant defense system in vivo has been elucidated previously. First, Zn can protect cell membranes against iron-initiated lipid oxidation by occupying negatively charged sites with potential iron binding capacity [9]. Second, Zn is the cofactor of antioxidant enzymes including Cu/Zn SOD and metallothioneins [7,28], which can serve as free radical scavengers. It is necessary to note that the most favorable antioxidant capacity in the liver and jejunum was observed when supplementing 60 mg/kg Zn instead of 80 mg/kg. While for ileum, only 20 mg/kg Zn from ZnPal increased its Cu/Zn SOD activity, and the response of SOD activity to Zn supplementation in the serum was lacking, despite the fact that serum MDA concentration was linearly reduced with the increasing level of ZnPal in the diet. However, a clear explanation cannot be elucidated based on our knowledge of broilers at the present study, and these different responses of SOD activity to dietary Zn level may result from Zn deposition capacity of tissues, metabolic activity, amount and speed of ROS generation, and distribution of SOD enzymes.
In conclusion, feeding diets supplemented with 0, 20, 40, 60, and 80 mg/kg Zn in the form of ZnPal did not affect the growth performance of broilers (weight gain, feed intake, and feed conversion ratio) in the 21-day study. Dietary ZnPal supplementation can promote the accumulations of hepatic trace mineral elements (Zn and Mg) and prevent lipid peroxidation by inhibiting serum MDA generation, with the optimal dosage of Zn from ZnPal being 80 mg/kg diet (analyzed Zn content in diet: 165 mg/kg), and it appeared that 60 mg/kg Zn as ZnPal (analyzed Zn content in diet: 148 mg/kg) was the optimum dosage for broilers to achieve maximum antioxidant enzyme activities.
ACKNOWLEDGMENTS
The technical assistance of our labmates are gratefully acknowledged.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.