Genetic Effects of Polymorphisms in Myogenic Regulatory Factors on Chicken Muscle Fiber Traits
Article information
Abstract
The myogenic regulatory factors is a family of transcription factors that play a key role in the development of skeletal muscle fibers, which are the main factors to affect the meat taste and texture. In the present study, we performed candidate gene analysis to identify single-nucleotide polymorphisms in the MyoD, Myf5, MyoG, and Mrf4 genes using polymerase chain reaction-single strand conformation polymorphism in 360 Erlang Mountain Chickens from three different housing systems (cage, pen, and free-range). The general linear model procedure was used to estimate the statistical significance of association between combined genotypes and muscle fiber traits of chickens. Two polymorphisms (g.39928301T>G and g.11579368C>T) were detected in the Mrf4 and MyoD gene, respectively. The diameters of thigh and pectoralis muscle fibers were higher in birds with the combined genotypes of GG-TT and TT-CT (p<0.05). Moreover, the interaction between housing system and combined genotypes has no significant effect on the traits of muscle fiber (p>0.05). Our findings suggest that the combined genotypes of TT-CT and GG-TT might be advantageous for muscle fiber traits, and could be the potential genetic markers for breeding program in Erlang Mountain Chickens.
INTRODUCTION
Because of relatively lower content of fat and cholesterol, and higher level of long-chain polyunsaturated fatty acids, many nutritionists believe that the white meat like poultry, seafood and fish is judged healthier than red meat, such as pork, beef and mutton. Combined with the lower price and rare religious restrictions, the consumption demand of chicken is increasing gradually (Yin et al., 2013). Nowadays, with the improvement of life level and the development of social economy, people begin to lay stress on food quality (Wang et al., 2009). The requirement of free-range chicken is much popular, because it is considered to be more healthier and more suitable for traditional eating habits that require the meat has higher chewiness, especially in southwest China (Dou, 2009). Previous studies have shown that the muscle fiber traits are the main factors to affect the chicken taste and texture (Branciari et al., 2009). The muscle fiber traits are regulated by several genes, the traditional phenotypic selection is very difficult to achieve the genetic improvement in a short time. However, the candidate gene approach is a powerful method to find the quantitative trait loci (QTL) for improving the rate of genetic progress in the meat quality traits (Yin et al., 2011).
In 1987, MyoD was first discovered by subtractive hybridization using myoblast cDNA libraries, which is a basic helix-loop-helix transcription factor that has ability to transform non-muscle cells, such as fibroblasts, into cells that are able to fuse into myotubes (Davis et al., 1987). Subsequently, three related skeletal muscle specific transcriptional factors—Myf5, myogenin (MyoG), and Mrf4—which are also capable of inducing myoblast features in non-muscle cell lines, were identified (Braun et al., 1989; Edmondson and Olson, 1989; Braun et al., 1990). Collectively, MyoD, Myf5, MyoG, and Mrf4 are specifically expressed in skeletal muscle linage and are therefore referred to as myogenic regulatory factors (MRFs) (Weintraub et al., 1991). Proteins of MRFs share a highly conserved basic helix-loop-helix domain that mediates DNA binding. Helix-loop-helix motif is essential for heterodimerization with a ubiquitously expressed E-protein (Massari and Murre, 2000). In the last few decades, studies have made significant advances in distinguishing the function of individual MRFs in myogenesis. It is shown that MyoD and Myf5 are factors involved in determination of myogenic cells, whereas MyoG and Mrf4 are more likely associated with terminal differentiation and homeostasis of myofibers (Singh and Dilworth, 2013).
Due to the very important roles of MRFs in control of the myogenic process, the genetic difference of the MRFs could affect the muscle fibers. To verify this hypothesis, we investigated the genetic polymorphisms of the MyoD, Myf5, MyoG, and Mrf4 genes in chickens and analyzed their association with muscle fibers traits.
MATERIALS AND METHODS
Chicken populations
The line SD02×SD03 of Erlang mountainous chicken was employed in the current study, which is a breeding population by Sichuan Agricultural University. From 1 to 42 d, 2,000 chicks with sexes intermingled were reared in concrete floor pens with shavings as litter. At d43, 1,200 chickens with similar body weight (314.6±24.1 g) were then randomly assigned to 3 different housing systems (400 birds/per group), including conventional cages that contained plastic hollowed pads, indoor floor pens with paddock, and outdoor free-range. Ten chickens were reared in each of 20 cages (1.9×1.0×0.7 m; 5.3 birds/m2). Indoor concrete floor pens were covered with wood shavings (3×5×2.7 m, 3.3 birds/m2), and chickens in this housing system had access to the paddock. The area ratio of indoor to paddock was 1:1. Chickens in the free-range treatment were raised under pines (6,670 m2) with a simple house (6.7 birds/m2) for shelter, and chickens were herded to pine grove for exercise in the daytime. Chickens in different systems were fed the same diet. Feed and tap water were freely available, and all diets were formulated to the feeding standards for yellow-feather chickens in China (NY/T33, 2004).
Sample collection and trait measurement
Blood samples were collected from the brachial vein in another 360 chickens (120 each randomly selected from three housing systems at d84, and the ratio of male and female was 1:1). Genomic DNA was extracted by the standard phenol-chloroform method.
The diameter and density of the muscle fibers were measured on the left pectoralis major muscle and thigh muscle of all selected chickens. The muscle slices (3 cm×2 cm) were cut to follow the direction of muscle fibers in the center of the muscle, then fixed overnight with 10% formalin. Fixed tissues embedded in paraffin were sectioned at 6 μm mounted on slides and stained with hematoxylin-eosin. Fiber diameter measurements were performed on each section under light-microscopy (40×) and the number of myofibers per square millimeter was computed.
Single-nucleotide polymorphism identification
Genetic variants in the MRFs genomic DNA sequences were analyzed using the polymerase chain reaction (PCR)-single strand conformation polymorphism method (SSCP). Twenty primer pairs were designed according to the Gallus gallus MRF genes sequences (Table 1) to amplify the eleven exons fully containing the coding sequences of the four genes. The PCR reactions included 0.8 μL (100 ng/μL) of template DNA, 5 μL of 2×Taq PCR MasterMix (TianGen Biochemical Technology, Shanghai, China), 0.4 μL of each primer (10 pmol/μL), and ddH2O to 10 μL reaction volume using the following condition: 95°C for 10 min, 35 cycles of 94°C for 30 s, annealing from 57°C to 62°C for 45 s, 72°C for 1 min, and a final extension at 72°C for 10 min. After PCR, single-nucleotide polymorphism (SNP) analysis was performed according to Orita et al. (1989). Six DNA samples showing identical patterns on the SSCP gel were further amplified in a 50 μL reaction volume, electrophoresed and purified with a gel extraction kit (Takara, Dalian, China), and sequenced by Shanghai Yingjun Biology Technique Corporation. The primer pairs used to amplify the fragments were also used for sequencing.
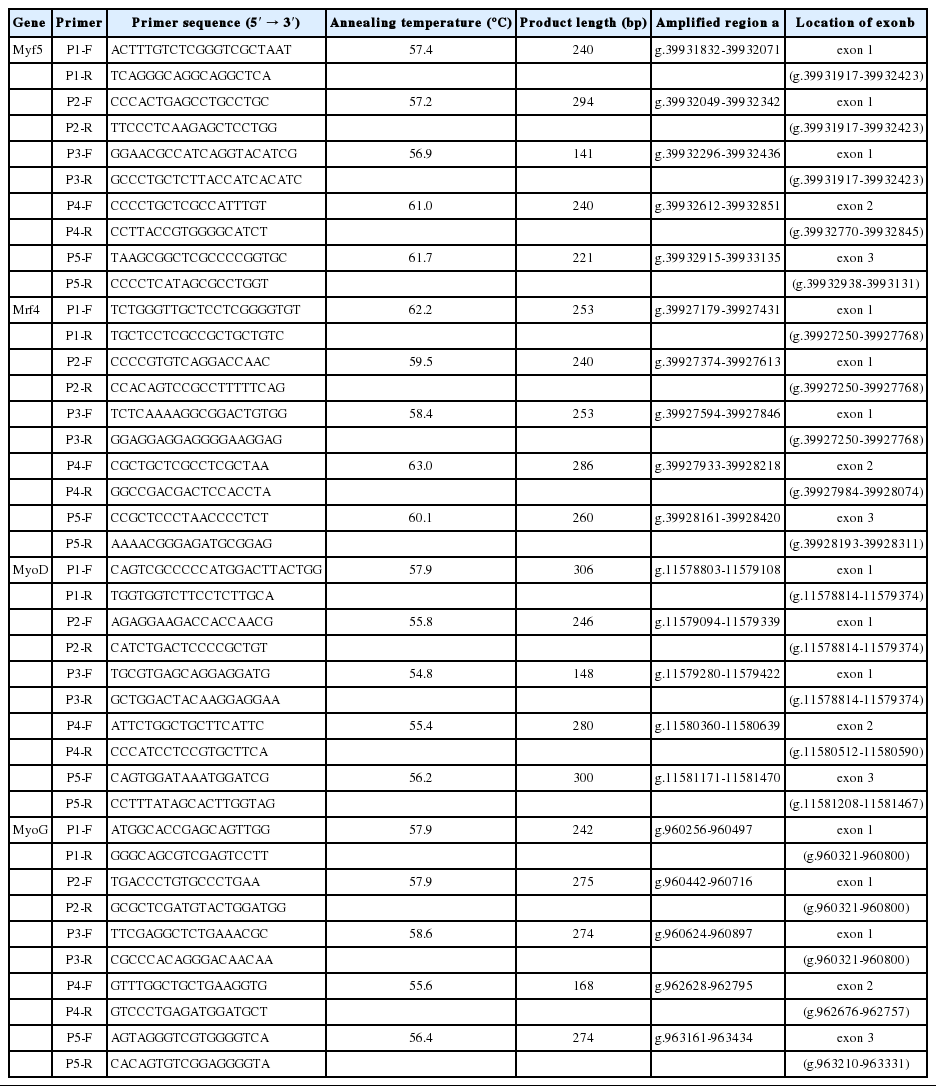
Primers for SNP detection
Data analysis
Combined genotypes were constructed based on the detected SNPs in all 360 experimental chickens. Genetic effect of the combined genotype was evaluated with the following model:
where Yijk is the trait measured in chicken; μ is the population means of that trait; Ci is the fixed effect associated with the combined genotypes i; Hj is the fixed effect associated with the housing system j, Sk is the fixed effect of sex k; eijk is the residual random effect, and the C×H, H×S, C×S, and C×H×S is the interaction effect of combined genotype i by housing system j, housing system j by sex k, combined genotype i by sex k, and combined genotype i by housing system j by sex k, respectively. Multiple comparisons were carried out using Tukey’s test. Pearson’s chi-square test was used to check for Hardy-Weinberg equilibrium of the SNP detected in the sample population. Polymorphism information content (PIC) was evaluated by Botstein’s method (Orita et al., 1989).
Data were presented as least square means±standard error of the mean, and differences were considered significant at p<0.05.
RESULTS
Single-nucleotide polymorphism identification and genetic diversity analysis
Twenty expected fragments (five fragments per gene) covering the full length of the coding sequences of Myf5, MyoD, Mrf4, and MyoG were amplified by the primers. Among them, two polymorphisms were detected by directly sequencing the polymorphic fragment based on the SSCP banding patterns, one of which was g.39928301T>G in the Mrf4 gene (Figure 1), and another is the mutation of g.11579368C>T in the MyoD gene (Figure 2). The variant of g.39928301T>G was a non-synonymous mutation and caused a GTG (Histidine) to GGG (Proline) change, while g.11579368C>T was a synonymous mutation that was not associated with an amino acid substitution.

The partial sequencing result of Mrf4 gene, and g. 39928301T>G was marked by rectangle.
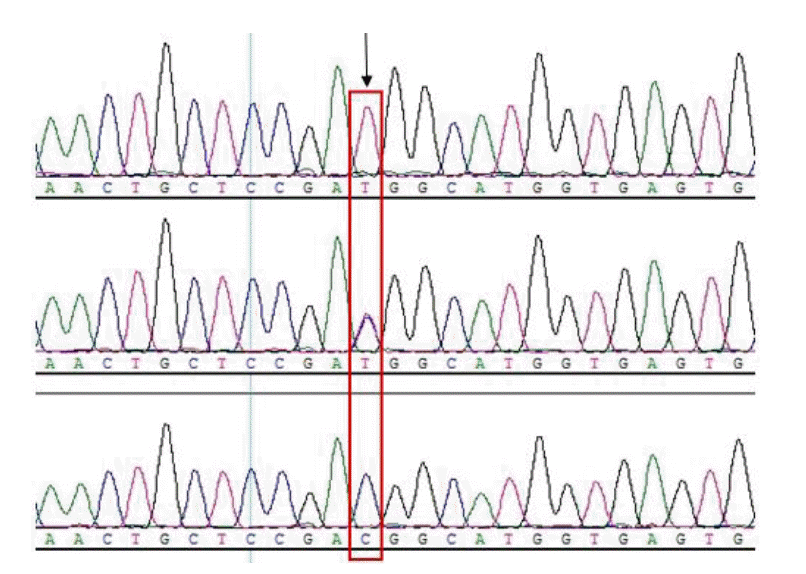
The partial sequencing result of MyoD gene, and g.11579368C>T was marked by rectangle.
Two SNPs, their corresponding allele and genotype frequencies, PIC, and p-values for the Hardy-Weinberg equilibrium test are shown in Table 2. Three genotypes each from two SNPs were identified. Genotypes TT, TG, and GG of g.39928301T>G had an average frequency of 42.22%, 35.56%, and 22.22%; while genotypes CC, CT, and TT of g.11579368C>T with frequencies of 46.11%, 32.78%, and 21.11%, respectively. The allele T was the dominant allele due to the higher frequency (60.00%) compared to the allele G (40.00%) in the SNP of g.39928301T>G. For SNP-g.11579368C>T the allele C represented the higher allele frequency (62.50%). Both g.39928301T>G and g.11579368C>T were out of Hardy-Weinberg equilibrium (p<0.05), and the PIC of them was 0.3648, and 0.3593, respectively.
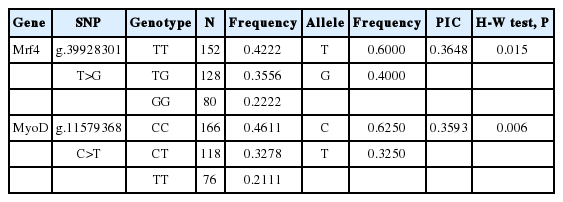
Frequency of alleles and genotypes, PIC, and the Hardy-Weinberg (H-W) equilibrium test at polymorphic sites
Effect of combined genotypes on the muscle fiber traits
We constructed the combined genotypes from each genotypes of g.39928301T>G and g.11579368C>T (Table 3). In the total 360 samples, there are 9 combined genotypes, those combined genotypes with low frequencies (TT-TT [n = 13, 3.61%], TG-TT [n = 13, 3.61%], GG-CC [n = 13, 3.61%], and GG-CT [n = 17, 4.72%]) were not included for further analysis. Association of the combined genotypes of Myf4 and MyoD with muscle fiber traits is shown in Table 3. The diameters of thigh and pectoralis muscle fibers were higher in birds with the combined genotypes of GG-TT and TT-CT. The density of pectoralis muscle fibers in individuals with GG-TT was lower than TG-CC birds, but the five evaluated combined genotypes had similar densities of thigh muscle fibers. Additionally, there was no interaction of housing system and combined genotype on the traits of muscle fiber (p>0.05).
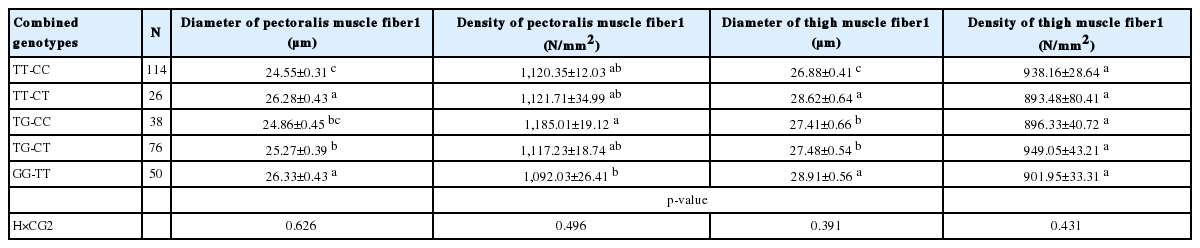
Association of the combined genotypes with muscle fiber traits in chickens
DISCUSSION
The diameter and density of muscle fiber directly determines the taste of meat, and thereby affects market acceptance. Although more chewy taste in chicken can be achieved through changing the housing system, the feed conversion ratio and growth performances are concurrently decrease (Massari and Murre, 2000; Fanatico et al., 2008; Dou, 2009). Fortunately, selecting individuals with additional information on their genotype for makers associated with QTLs and target traits can effectively balance muscle fiber trait and growth performance in different housing systems. Therefore, we investigated the polymorphisms in the coding regions of Myf5, Mrf4, MyoD, and MyoG, and explored their association with muscle fiber traits. In the present study, two SNPs were detected, locating in exon 3 of the Mrf4 gene, and in exon 1 of the MyoD gene. The PIC used to assess the genetic variation, is divided into three classes, including slight variation (PIC< 0.25), medium polymorphic (0.25<PIC<0.5), and highly informative (PIC>0.5) (Yin et al., 2013). The PIC of 0.3648 and 0.3593 in two SNPs indicated that this population has higher selective space for breeding.
Although the function of synonymous mutations remain unknown, recent studies revealed that silent SNPs can lead to the synthesis of protein products with the same amino acid sequence but different structural and functional properties (Komar, 2007), and alter the structure of substrate and inhibitor interaction sites (Kimchi-Sarfaty et al., 2007). Therefore, we presumed that the synonymous mutation of g.11579368C>T in the MyoD gene changes the style of protein folding or it may link to another mutation of g.39928301T>G in the coding regions of the Mrf4 gene that influence the muscle fiber traits. In the present study, the results showed that the combined genotypes of mutations in MyoD and Mrf4 are associated with variations in the muscle fiber traits. The combined genotypes of TT-CT and GG-TT with greater pectoralis and thigh muscle fiber diameters, and could be selected in a breeding program to improvement of the chicken muscle fiber characteristics.
Environmental sensitivity is the ability of a genotype to alter its phenotypic expression in response to environmental influences. The genetic variation in environmental sensitivity is used as a definition of genotype by environment interaction (Kolmodin et al., 2003). There is a reranking of individuals for traits expression because of the varying magnitude of phenotypic change in the different environments. Consequently, the selection for target performance in one environment may not result in enhanced performance under other environmental conditions (Sing, 1990). In the present study, we screened the polymorphisms in the coding sequences of MRFs of chickens from three housing environments, and analyzed their relationship with muscle fiber traits. Fortunately, we found that there was no interaction between housing system and combined genotypes, which showed that the advantageous combined genotypes of TT-CT and GG-TT may promote the diameter of muscle fiber under different rearing conditions.
CONCLUSION
In conclusion, the present study showed that the combined genotypes of TT-CT and GG-TT may serve as molecular makers for breeding program. Although we can use marker-assisted selection to increase muscle fiber diameter, a greater diameter does not necessarily equate to superior meat quality. Thus, we should find the balance between the diameter of the muscle fiber and meat quality in the further studies.
ACKNOWLEDGMENTS
This work was supported by the China Agriculture Research System (CARS-41), the Twelfth Five Year Plan for breeding programs in Sichuan—Selective breeding of new breeds and the synthetic strains in laying hens (2011NZ0099-7).