Effects of Heat-oxidized Soy Protein Isolate on Growth Performance and Digestive Function of Broiler Chickens at Early Age
Article information
Abstract
This study was conducted to investigate effects of heat-oxidized soy protein isolate (HSPI) on growth performance, serum biochemical indices, apparent nutrient digestibility and digestive function of broiler chickens. A total of 320 1-day-old Arbor Acres chicks were randomly divided into 4 groups with 8 replicates of 10 birds, receiving diets containing soy protein isolate (SPI, control group) or the same amount of SPI heated in an oven at 100°C for 1, 4, or 8 h, for 21 days, respectively. The results indicated that compared with the control group, body weight gain and feed intake of birds fed diet containing SPI heated for 8 h were significantly lower (p<0.05). Serum urea nitrogen concentration was higher in the broilers fed diet containing SPI heated for 4 or 8 h at d 21 (p<0.05). In contrast, serum glucose content was decreased by HSPI substitution at d 21 (p<0.05). The relative pancreas weight in HSPI groups was higher than that in the control group at d 21 (p<0.05). Meanwhile, the opposite effect was observed for relative weight of anterior intestine and ileum in broilers fed a diet containing SPI heated for 8 h (p<0.05). Birds fed diets containing SPI heated for 4 or 8 h had a decreased lipase activity in anterior intestinal content at d 14 and 21 (p<0.05), respectively. In addition, the same effect was also noted in broilers given diets containing SPI heated for 1 h at d 21 (p<0.05). Similarly, amylase, protease and trypsin activity in anterior intestinal content were lower in broilers fed diets containing SPI heated for 8 h at d 21 (p<0.05). The apparent digestibility of dry matter (DM) from d 8 to 10 and DM, crude protein (CP), and ether extract from d 15 to 17 were lower in broilers fed diets containing SPI heated for 8 h (p<0.05). Besides, birds given diets containing SPI heated for 4 h also exhibited lower CP apparent digestibility from d 15 to 17 (p<0.05). It was concluded that HSPI inclusion can exert a negative influence on the growth performance of broilers, which was likely to result from the simultaneously compromised digestive function.
INTRODUCTION
Protein has been known to be the most important nutrient in animal feed, providing not only amino acids for body protein synthesis, but also energy for growth (Korhonen et al., 1998). Protein quality plays vital role in protein digestibility and utilization. Protein oxidation is the structural modification induced directly by reactive oxygen species or indirectly by reaction with the by-products of lipid peroxidation (Shacter, 2000). During processing and storage, food protein can be easily affected by various factors, such as heat, comminuting and grinding, which introduce molecular oxygen and mix oxidants with susceptible food components (Korhonen et al., 1998). Oxidative modification of proteins has been shown to induce a number of multiple structural changes such as oxidation of side chains of special amino acid residues and peptide chains in protein, fragmentation, aggregation, and changes in hydrophobicity as well as conformation (Headlam and Davies, 2004). Ultimately, these alterations often result in changes in functional properties and bioavailability of protein (Witko-Sarsat et al., 2003; Smet et al., 2008; Zhang et al., 2011). Studies have claimed that oxidative modification of protein can lead to altered digestibility of proteins in vitro and susceptibility of protein to protein-degrading enzymes. Zamora and Hidalgo (2001) reported that proteolytic susceptibility of proteins to chymotrypsin, pancreatin, pronase and trypsin was decreased by oxidative modification induced by lipid peroxidation. Similarly, Sante-Lhoutellier et al. (2007) found a direct and quantitative relationship between oxidized myofibrillar proteins caused by hydroxyl radical and decreased proteolytic susceptibility to proteases of the digestive tract. Moreover, it was reported that heat oxidation led the soy protein to generate reactive oxygen species, decrease the antioxidant status and induce redox imbalance in digestive organs of mice fed heat-oxidized soybean protein (Tang et al., 2012a). Besides, heat-oxidized soy protein isolate (HSPI) also caused compromised growth performance and inferior immune function of broilers (Wu et al., 2014). These studies provide evidences that oxidative modification may adversely affect protein digestibility and digestive function of animals. Nevertheless, few studies have been carried out concerning the effects of dietary oxidized protein on growth performance and digestive function of broilers.
Heating is one of the most common and widely used methods in many different food products to modify proteins (Tang et al., 2012b; Wu et al., 2014). Soy protein isolate (SPI) is used as a functional ingredient in many different food products and it has been reported that SPI is usually subjected to heating during processing (Renkema et al., 2002; Tang et al., 2009; Wu et al., 2014). Therefore, SPI heated in an oven at 100°C for 1, 4, and 8 h, respectively, were used in this study and the aim was to determine whether dietary oxidized protein would have any negative effects on the growth performance and digestive function of broilers at early age.
MATERIAL AND METHODS
Materials
SPI was purchased from Sun-Green Biotech Co., Ltd. (Nantong, China). Analyzed moisture, crude protein (CP), ether extract (EE) and ash contents in the SPI were 5.6%, 86.2%, 0.4% and 4.9%, respectively. The HSPI was prepared according to the method as described by Tang et al. (2012a, b) and Wu et al. (2014). 1.0 kg native SPI was evenly placed on clean and dry stainless steel tray (Length×Width×Height:14 cm×8 cm×2 cm) and was dry heated at 100°C (1, 4, 8 h), then air-dried under normal condition (around 25°C) and stored at 4°C. Protein carbonyl content in SPI heated for 4 h (17.80 nmol/mg prot), and 8 h (22.35 nmol/mg prot) were significantly higher than normal SPI (15.13 nmol/mgp rot) and SPI heated for 1 h (15.79 nmol/mg prot).
Experimental design, diets and management
All experimental conditions and animal procedures were approved by Nanjing Agricultural University Institutional Animal Care and Use Committee. A total of 320 1 day-old Arbor Acres broiler chicks (45.01±0.07 g) from a commercial hatchery (Hewei Co., Ltd, Anhui, China) were used in this study. The birds were randomly divided into 4 groups with 8 replicates of 10 birds and were fed a diet containing normal SPI (Control) or diets containing the same amount of SPI heated in an oven at 100°C for 1, 4, and 8 h, respectively. The experiment lasted for 21 d. The basal diet was formulated based on the NRC (1994) to meet the nutrient requirements of the broiler. The formulation and nutrient level of basal diet were shown in Table 1.

The formulation and calculated nutrient levels of basal diet (%, as-fed basis unless otherwise stated)
Birds were housed in an environmentally controlled room. Temperature was maintained at 32°C for the first week and then gradually reduced, according to normal management practices, until a temperature of 26°C was achieved. Continuous light was provided throughout the entire experiment. Mash feed and fresh water were available ad libitum at all growth phase. Body weight and feed intake during the 21-d experiment (measured at 1 and 21 d after deprivation of feed for 12 h, with water being provided ad libitum) were recorded to calculate body weight gain, feed intake and feed conversion rate.
Sample collection
On d 14 and 21, 8 birds from each treatment (one bird per cage) were randomly selected and weighed after feed deprivation for 12 h. Blood samples were collected from the wing vein. Serum samples were then taken by centrifugation at 3,000×g for 15 min at 4°C and stored at −20°C until analysis (Chen et al., 2013). After blood sampling, birds were killed by cervical dislocation immediately. The proventriculus, gizzard, pancreas, duodenum (from ventriculus to pancreo-biliary ducts), jejunum (from pancreo-biliary ducts to yolk stalk) and ileum (from yolk stalk to ileocecal junction) were removed and weighed (without content and gall bladder). Relative organ weight was calculated according to following formula (Wu et al., 2014): relative weight of digestive organs (g per 100 g body weight) = organ weight (g)/body weight (100 g). After that, contents within anterior intestine (duodenum and jejunum) were collected by massaging the tract from both ends into plastic containers and stored at −20°C until analysis.
Serum biochemistry
Serum urea nitrogen (UN, berthelot color reaction method), total protein (TP, biuret method), albumin (ALB, bromocresol green method), triglyceride (TG, glycerol kinase method), total cholesterol (TC, cholesterol esterase/peroxidase method) and glucose (GLU, glucose oxidase method) were determined on a biochemical autoanalyzer (Beckman Instruments, Inc. Scientific Instruments Division, Fullerton, CA, USA) according to the manufacturer’s recommended procedure (Fossati and Prencipe, 1982; Thomas et al., 1982; Deeg and Ziegenhorn, 1983; Denli et al., 2009).
Digestive enzyme assay
Anterior intestinal content samples were homogenized (1:9, wt/vol) with ice-cold 154 mmol/L sodium chloride solution, and then centrifuged at 4,550×g for 15 min at 4°C. The supernatants were divided into small portions and stored for enzyme activity assay. Enzymes activities were determined according to method used in our lab as described by Wen et al. (2012). In detail, enterokinase was added to homogenate to activate pancreatic zymogens before determination of pancreatic trypsin and protease. Amylase activity was determined using the iodometric method and one amylase unit was defined as the amount of enzyme that hydrolyzed 10 mg of starch in 30 min. Protease activity was assayed using casein as substrate and reacting it with Folin reagent. One protease unit was defined as the amount of enzyme that hydrolyzed casein to form l μmol product per min. Trypsin activity was assessed using N-benzoyl-l-arginine ethyl ester as substrate. One unit of trypsin was defined as the amount of enzyme that increased 0.003 of absorbance of product per minute. Lipase activity was determined using a turbidimetric method and one lipase unit was defined as the amount of enzyme that hydrolyzed l μmol of olive oil per min. Protein concentration was determined using bovine ALB as a standard. Amylase and protease activities were expressed as units per milligram of protein, trypsin activity was expressed as units per microgram of protein, and lipase activity was expressed as units per gram of protein.
Digestibility determination
During days 8 to 10 and 15 to 17, uncontaminated excreta (free from feathers and feed) were collected twice per day (0800 and 1600 h) from each cage. The excreta samples from the three collections from each cage were pooled and then dried for 48 h in an oven at 65°C. The dried excreta were allowed to equilibrate to atmospheric conditions for 24 h before being weighed. Feed and excreta samples were then ground through a 0.45-mm screen and subsequently analyzed (AOAC, 1990) for dry matter (DM, 934.01), EE (920.39) and CP (954.01). Apparent digestibility was determined using acid-insoluble ash (Acid-insoluble, 920.72) as the indicator and was expressed as %.
Statistical analysis
All data were analyzed as a completely randomized design using one-way analysis of variance with Duncan multiple comparison test (SPSS, 2008). The differences were considered to be significant at p<0.05. The p values between 0.05 and 0.10 were considered as a trend.
RESULTS
Growth performance
As expected (Table 2), the body weight and feed intake were lower in HSPI groups especially the SPI heated for 8 h group (p<0.05) when compared with the control. However, feed conversion ratio was not affected by HSPI (p>0.05).
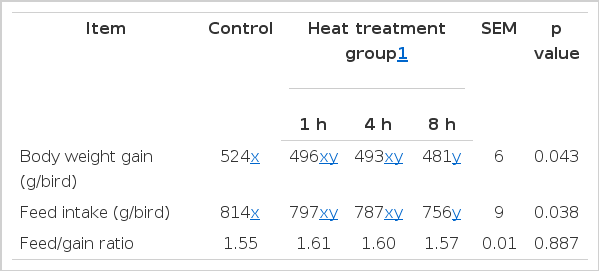
Effects of heat-oxidized soy protein isolate on growth performance of broilers
Serum biochemical indices
Dietary HSPI can cause an increase in serum UN content and this negative effect was more pronounced in long-term heat-oxidized SPI groups as demonstrated by significantly increased UN content in broilers fed diets containing SPI heated for 4 h (0.30 vs 0.36 mmol/L) or 8 h (0.30 vs 0.40 mmol/L) at d 21 (p<0.05) when compared with the control (Table 3). In contrast, dietary HSPI inclusion significantly decreased serum GLU content at d 21 (p<0.05). Other serum biochemical indices (TC, TG, TP, and ALB) were unaffected by HSPI substitution (p>0.05).

Effects of heat-oxidized soy protein isolate on serum biochemical indices of broilers
Organs weights
As summarized in Table 4, no significant differences were noticed for the relative proventriculus or gizzard weight among groups at both d 14 and 21 (p>0.05). Compared with the control group, dietary HSPI resulted in a significant increase in the relative pancreas weight at d 21 (p<0.05). In contrast, relative anterior intestine and ileum weight were significantly decreased by SPI heated for 8 h at d 21 (p<0.05), but this effect was not observed in the other HSPI groups (p>0.05).

Effects of heat-oxidized soy protein isolate on relative weight of digestive organs (g/100 g body weight) of broilers
Digestive enzyme activity
There were no significant (Table 5) differences in amylase, protease, and trypsin activity among groups at d 14 (p>0.05), however, the activities of those enzymes were significantly decreased by SPI heated for 8 h as compared with control group at d 21 (p<0.05). As for the lipase, broilers fed diets containing SPI heated for 4 h or 8 h exhibited a significantly lower activity than those in control group at either d 14 or 21 (p<0.05), and the same effect was also observed in broilers given SPI heated for 1 h at d 21 (p<0.05).
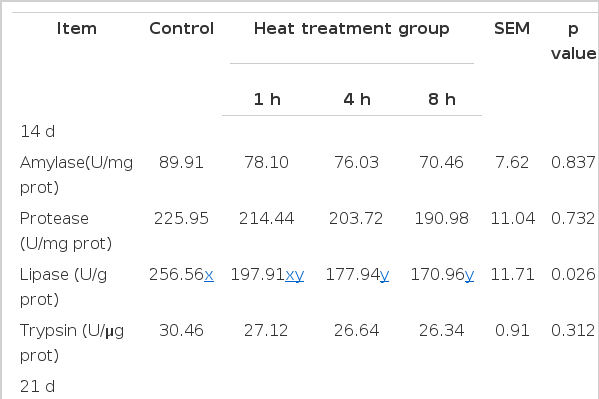
Effects of heat-oxidized soy protein isolate on digestive enzyme activities within anterior intestinal contents of broilers
Apparent digestibility of nutrients
HSPI substitution (Table 6) did not affect the apparent digestibility of CP and EE from d 8 to 10 (p>0.05). As expected, compared with control, the apparent digestibility of DM of birds from d 8 to 10 and DM, CP and EE from d 15 to 17 were significantly decreased by SPI heated for 8 h (p<0.05). Besides, birds given diet containing SPI heated for 4 h also exhibited lower CP apparent digestibility from d 15 to 17 (p<0.05).
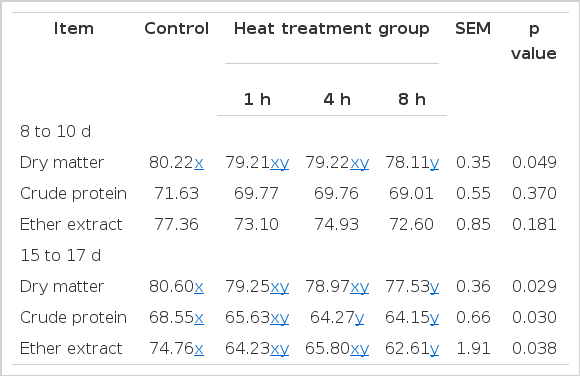
Effects of heat-oxidized soy protein isolate on apparent nutrient digestibility of broilers (%)
DISCUSSION
Our study has shown that HSPI inclusion significantly decreased body weight gain of broilers, which was likely the result from simultaneously compromised feed intake rather than similar feed conversion ratio. Similar result was also observed by Tang et al. (2012a) in mice and in broilers by Wu et al. (2014). The inferior growth performance was due to inferior digestive function discussed in the following.
Serum biochemistry is a labile biochemical system which can reflect the condition of the organism and the changes happening to it under influence of internal and external factors. In the current study, broilers fed diets containing HSPI exhibited lower serum GLU level. The decrease in the serum GLU level was quite interesting because birds generally maintain a high and relatively constant serum GLU level even in low feed intake (Liukkonen-Anttila, 2001). We would likely to attribute the lower GLU content to the inferior nutrient digestion capacity of birds fed HSPI diets. Changes in serum UN concentration can reflect the whole body status of amino acid metabolism and utilization in animals. Increased accretion of serum UN concentration would suggest a potential attenuation of synthesis of protein in animals. Previous studies have indicated that oxidative modification adversely affects numerous amino acids in protein or peptides (Stadtman, 1990; Wu et al., 2014). Therefore, changes of amino acid composition in HSPI may be responsible for the deterioration of synthesis of protein in broilers. In the present study, serum UN of broilers was significantly higher in broilers fed diets containing SPI heated for 4 and 8 h, suggesting that oxidative modification may decline the protein quality and utilization of the diet.
The relative weight is one of the primary biological characteristics of digestive organs and the metabolic activity of digestive organs is a function of the metabolic activity and size of the organs. In the current study, the relative pancreas weight of broilers fed HSPI was significantly heavier than that of the control group at d 21, indicating that oxidized protein can probably cause compensatory hypertrophy of pancreas, which may be due to changes in pancreatic cellular hypertrophy and cell number induced by HSPI inclusion (Gu and Hu, 2010). In addition, Mahmood et al. (1997) reported that lower feed intake itself could induce an increase in the relative weight of the pancreas. Therefore, the pancreatic hypertrophy may also result from the lower feed intake in broilers fed diets containing HSPI. Effects of HSPI inclusion on relative weight of other digestive organs were quite different from pancreas. In the current study, relative anterior intestine and ileum weight showed a significantly decrease in broilers fed a diet containing SPI heated for 8 h at d 21. These combined changes indicate that oxidized protein may have a negative impact on the development digestive tracts and organs, and the impairment may be due to growth depression observed in broilers fed HSPI.
Digestive enzymes in the intestine of poultry are mainly synthesized by and secreted from the pancreas. However, they can only be activated in the gut and then play their expected roles. Therefore, changes of intestinal enzymes will illustrate the development of the digestive process. Rideau et al. (1983) observed that intestinal enzymes were located mainly in the duodenum or jejunum in the chicken. In the current study, we therefore selected digestive enzymes present in the anterior intestine (duodenum and jejunum) as an indicator of intestinal digestive enzymes activities. Our data showed that HSPI inclusion decreased the amylase, protease, lipase and trypsin activities. The lower small intestinal enzyme activities observed in birds fed HSPI may be responsible for the impairment of growth performance because it is well known that digestive enzymes play a rate-determining role in providing the substrates for growth. Recently, it has been demonstrated in many studies that oxidative modification is responsible for the protein aggregation and the consequent decrease of digestibility (Gatellier et al., 2009; Promeyrat et al., 2010). In the current study, the decrease in the apparent digestibility of DM, CP, and EE of broilers fed diets containing HSPI was consistent with the study of Tang et al. (2012a), who reported that ingestion of a HSPI diet resulted in a significant decrease in apparent digestibility of the DM and CP in mice. It may then be inferred that ingesting diets containing oxidized protein can cause a negative effect on nutrient digestibility of broilers.
In conclusion, our study indicated that ingesting diets containing HSPI exerted a negative influence on growth performance and digestive function of broiler chickens as indicated by various measured parameters. More studies are needed to explain the precise mechanism involved in the modification of feed protein, and the potential health risk in animals induced by oxidized protein inclusion.
ACKNOWLEDGMENTS
We gratefully thank our labmates in the College of Animal Science and Technology of Nanjing Agricultural University for their assistant in this study. This work was supported by the Postgraduate Education Innovation Project of Jiangsu province in China (CXLX13-289).