



INTRODUCTION
Food allergies have become increasingly prevalent during the past few decades. It was recently reported that up to 6% of young children and 4% of adults are affected by food allergies (Sicherer et al., 2006). Cumulative evidences have revealed that there was a close relationship between the development of allergic diseases and the imbalance of intestinal microbiota. Bj├Črkst├®n et al. (1999) showed that the allergic children harbored lower counts of lactobacilli and bifidobacteria, but higher counts of coliforms and Staphylococcus aureus, as compared to non-allergic children. Sj├Čgren et al. (2009) also suggested that children who developed allergy were significantly less colonization of lactobacilli and Bifidobacterium adolescentis during the first 2 months of age. Johansson et al. (2011) found that the colonization of infants gut by a group of lactobacilli reduces the risk of allergy at 5 years of age despite these children had allergic heredity. Therefore, the balance of gut microbiota plays an important role in the prevention of allergic diseases.
Recently, developments in mouse models of food allergy have provided the new tools to better understand this disease. Animal models of food allergy have been used to explore mechanisms of the development of sensitization to food proteins as well as immunologic mechanisms of adverse reactions to allergen re-exposure (Kweon et al., 2000; Forbes et al., 2008; Brandt et al., 2009). However, little information was available on the gut microbiota change during the development of allergy model. Furthermore, all these models were developed under barrier systems, such as SPF condition, which are expensive to establish and maintain. Therefore, in this study, we have used a mouse model of ovalbumin-induced allergic diarrhea according to Brant et al. (2003) under a sub-barrier system, which is in line with international standard and having low cost, and investigated the development of gut microbiota during the allergic diarrhea induction, aiming to gain further insight into the relationship between the gut microbiota and allergy development and provide basic knowledge for the prevention of food allergy.
MATERIAL AND METHODS
Animals and experimental design
Twenty four male BALB/c mice (18 to 22 g, 6 to 8 week old) obtained from Nanjing Jinling Hospital (Nanjing, China) were housed under sub-barrier system and allowed free access to food and water. Mice were randomly allocated to two groups, OVA-sensitized treatment group and sham-sensitized control group (n = 12/group), after one week of adaptive phase. All experimental protocols were approved by the Animal Studies Committee of Nanjing Jinling Hospital.
Sub-barrier system
Laminar flow cabinets and super-clean bench in the laboratory were used for animal feed and experimental manipulation respectively during the whole experiment. Water, cage and beddings were sterilized by autoclaving. Feed were sterilized by radiation. In addition, walls, floors and others in the laboratory were fully cleaned and disinfected according to the requirement of sterile room.
Induction of allergic diarrhea
Mice were sensitized and challenged as previously by Brant et al. (2003). Briefly, mice in treatment group were sensitized to OVA (grade v, Sigma) by intraperitoneal injection of 100 ╬╝g of OVA in alum on d 0 and d 14, while mice in control group were sham-sensitized using saline, followed by intragastric feeding of 50 mg of OVA in sterile phosphate buffer saline (PBS, 0.1 mol/L, pH 7.4) on days 28, 30, 32, 34, 36 and 38. Diarrhea was assessed by visually monitoring mice for up to one hour following intragastric challenge. Mice showing profuse liquid stool were recorded as developing diarrhea. All mice were sacrificed after the 6th intragastric challenge and samples were collected for further analysis.
Measurement of immunoglobulins
Serum were processed and frozen at ŌłÆ80┬░C until analysis. Serum OVA-specific IgG1 levels were determined by enzyme linked immunosorbent assay (ELISA). In brief, wells were coated with 100 ╬╝g/ml OVA and blocked with 5% skim milk in water (w/v). Plates were washed with 0.05% Tween-20 in PBS and serial dilutions of serum samples (diluted 1:100,000 and then serially diluted 1:2) were added. After one-hour incubation, plates were washed and horseradish peroxidase-conjugated anti-mouse IgG1 (1:1,000) (A90ŌĆō105P, Bethyl Laboratories, Inc) was added. Finally, 100 ╬╝l of o-phenylenediamine (0688, Amresco) substrate solution was added. Colorimetric reaction was stopped with 2 mol/L H2SO4 and the optical density (OD) was read at 492 nm within 10 min. Data represent mean┬▒SE of the serum dilution required to obtain OD = 0.6, because it is in the middle of the linear part of the curve. The level of total IgE in the serum was measured using an ELISA kit (E-90E, Immunology Consultants Laboratory, Inc). Measurements were performed according to the manufacturesŌĆÖ instructions.
Intestinal mast cell quantification
Jejunum segments were fixed in 4% paraformaldehyde (w/v) and paraffin embedded for section and histological assessment. Mast cells were detected by chloroacetate esterase (CAE) staining according to published protocols of mast cell detection (Beckstead et al., 1981). The 5 ╬╝m sections were stained for CAE-positive mucosal mast cells and lightly counterstained with hematoxylin. Five random mice per group and at least three random sections per mouse were analyzed. The numbers of stained cells in a high-power field area (├Ś200) were counted using Image-pro Plus 6.0 software.
DNA extraction and PCR amplification
Total DNA was extracted from each colonic content sample according to a bead-beating method using a mini-bead beater (Biospec Products, Bartlesville, OK) and followed by phenol-chloroform extraction (Zoetendal et al., 1998). DNA was then precipitated with ethanol and pellets were dissolved in 50 ╬╝l of TE. Primers U968-GC and L1401 (see Table 1 for all the primers used in this study) (N├╝bel et al., 1996) were used to amplify the V6 to V8 variable regions of the bacterial 16S rRNA gene. PCR amplification was performed using the following program: 94┬░C for 5 min, and 35 cycles of 94┬░C for 30 s, 56┬░C for 20 s, 68┬░C for 40 s and 68┬░C for a 7-min final extension.
DGGE analysis
PCR amplicons obtained from V6 to V8 regions of 16S rRNA genes were separated by DGGE according to the specifications of Muyzer et al. (1993) using a Dcode TM system (Bio-Rad, Hercules, CA, USA). DGGE was performed in 8% polyacrylamide gels containing 37.5:1 acrylamide-bisacrylamide and a denaturing gradient of 40% to 53%. Electrophoresis was initiated by pre-running for 10 min at 200 V and subsequently continued at a fixed voltage of 85 V for 12 h. Gels were stained with AgNO3 after completion of electrophoresis (Sanguinetti et al., 1994). DGGE gels were scanned using a GS-800 Calibrated Densitometer (Bio-Rad) and principle component analysis (PCA) was performed using SIMCAP+(12) software.
Real-time PCR quantification of total bacteria, Lactobacillus and Bifidobacterium
Real-time PCR was performed on an Applied Biosystems 7300 Real-time PCR System (Applied Biosystems, California, USA). A reaction mixture (20 ╬╝l) consisted of 10 ╬╝l of IQ SYBR Green Supermix (Bio-Rad, California, USA), 0.2 ╬╝M of each primer set and 1 ╬╝l of the template DNA. Standard curves were generated with 10-fold serial dilutions of 16S rRNA genes amplified from the respective target strains. The concentration of 16S rRNA gene abundance was plotted against the CT value. Universal primers, Bact1369 and Prok1492 (Suzuki et al., 2000), were used to estimate the total number of copies of the bacterial 16S rRNA gene in each sample. Lactobacillus and Bifidobacterium were quantified using the primers Ulac16S1F/Ulac16S1R (Khafipour et al., 2009) and g-Bifid-F/g-Bifid-R (Matsuki et al., 2004) respectively. Samples from five random mice per group were analyzed.
RESULTS
Induction of intestinal allergic symptom
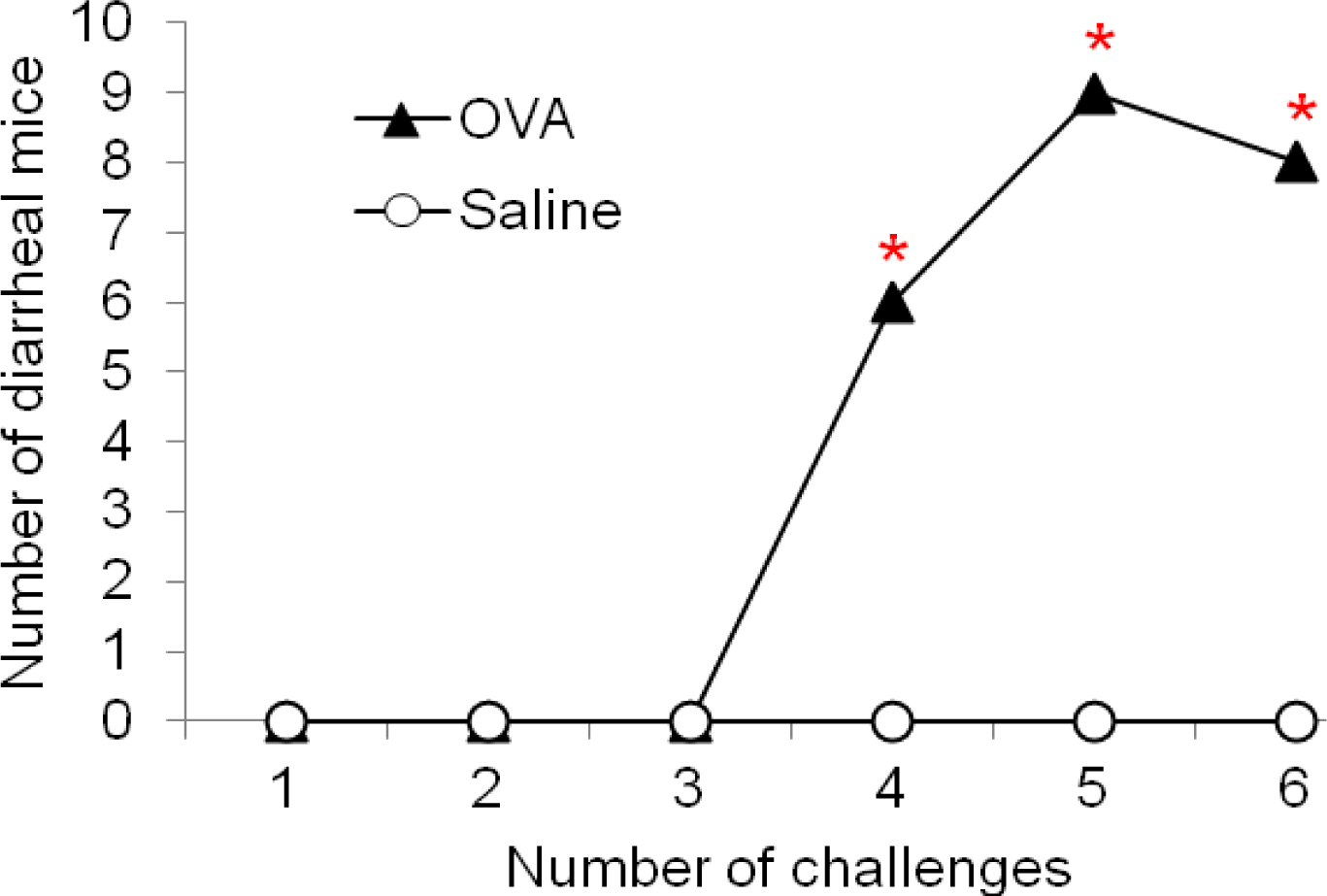
The stool of OVA-sensitized mice began to shift from normal to a profuse liquid stool after the 4th OVA challenge. In addition, the liquid stool was observed following OVA-induced diarrhea contrasts with the solid pellets seen in the distal colon of normal mice. Figure 1 shows the time course of changes in the number of mice affected with diarrhea. Diarrhea was observed after 4th oral administrations of OVA to the OVA-sensitized mice, and the incidence of diarrhea continued to increase by the end of the experiment. However, repeated oral administration of OVA in mice with sham-sensitization failed to induce any signs of diarrhea. In particular, OVA-sensitized mice showed a significantly higher incidence of diarrhea as compared with those without OVA-sensitization after 4th to 6th challenges (x2 test, p<0.05).
OVA-specific IgG1 and total IgE in the sera
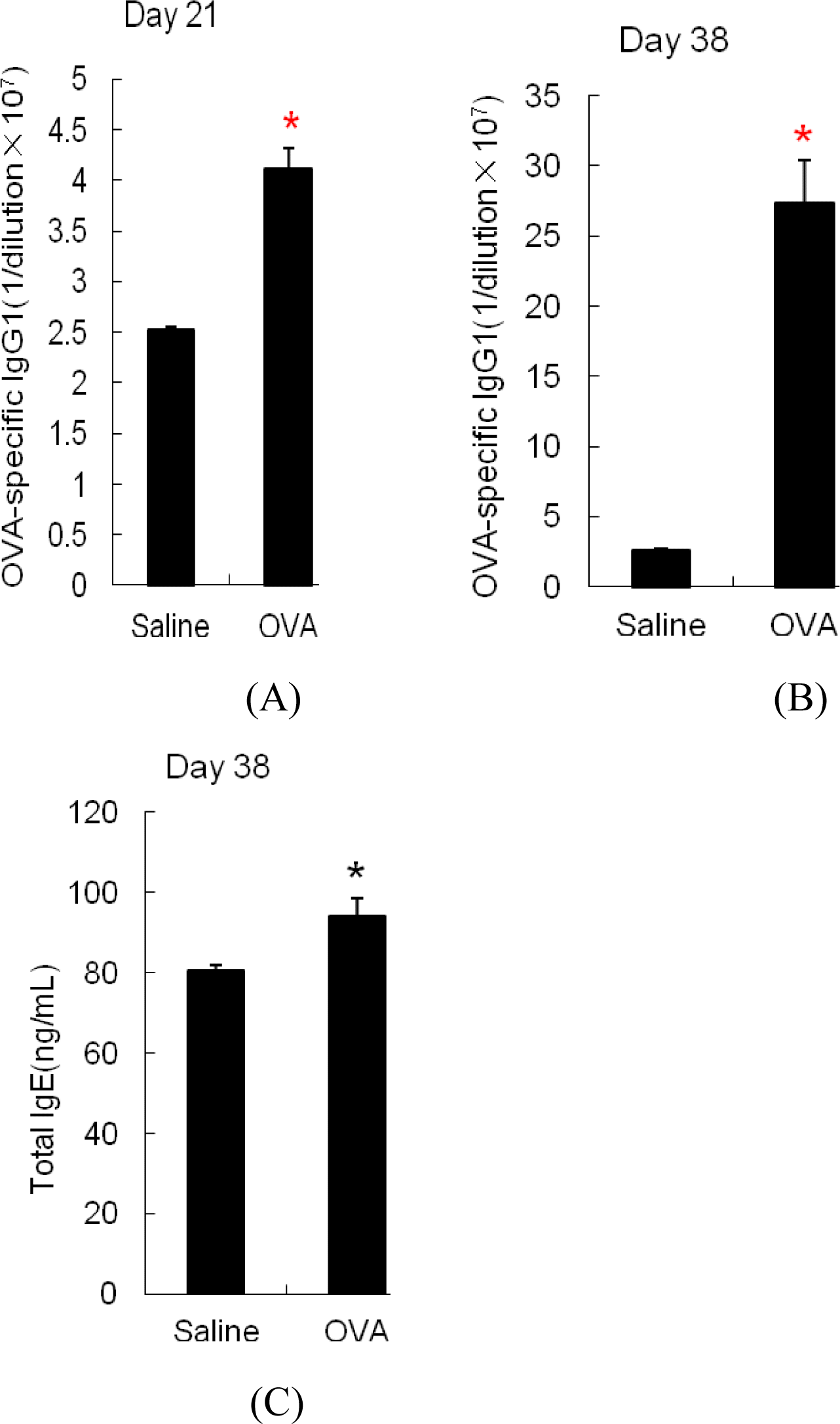
The level of OVA-specific IgG1 in the sera was measured on d 21 (one week after two sensitizations) and d 38 (after 6th challenge). As shown in Figure 2A, sera levels of OVA-specific IgG1 was significantly higher in OVA-sensitized mice compared with sham-sensitized mice after sensitization alone (p<0.05). Following allergen-induced diarrhea, OVA-specific IgG1 in OVA-sensitized mice had a further increase, which was approximately 10-fold greater than that of sham-sensitized mice (Figure 2B). In addition, total IgE sera titers after OVA challenges were significantly higher in OVA-sensitized mice compared with sham-sensitized mice (Figure 2C).
Mast cell infiltration
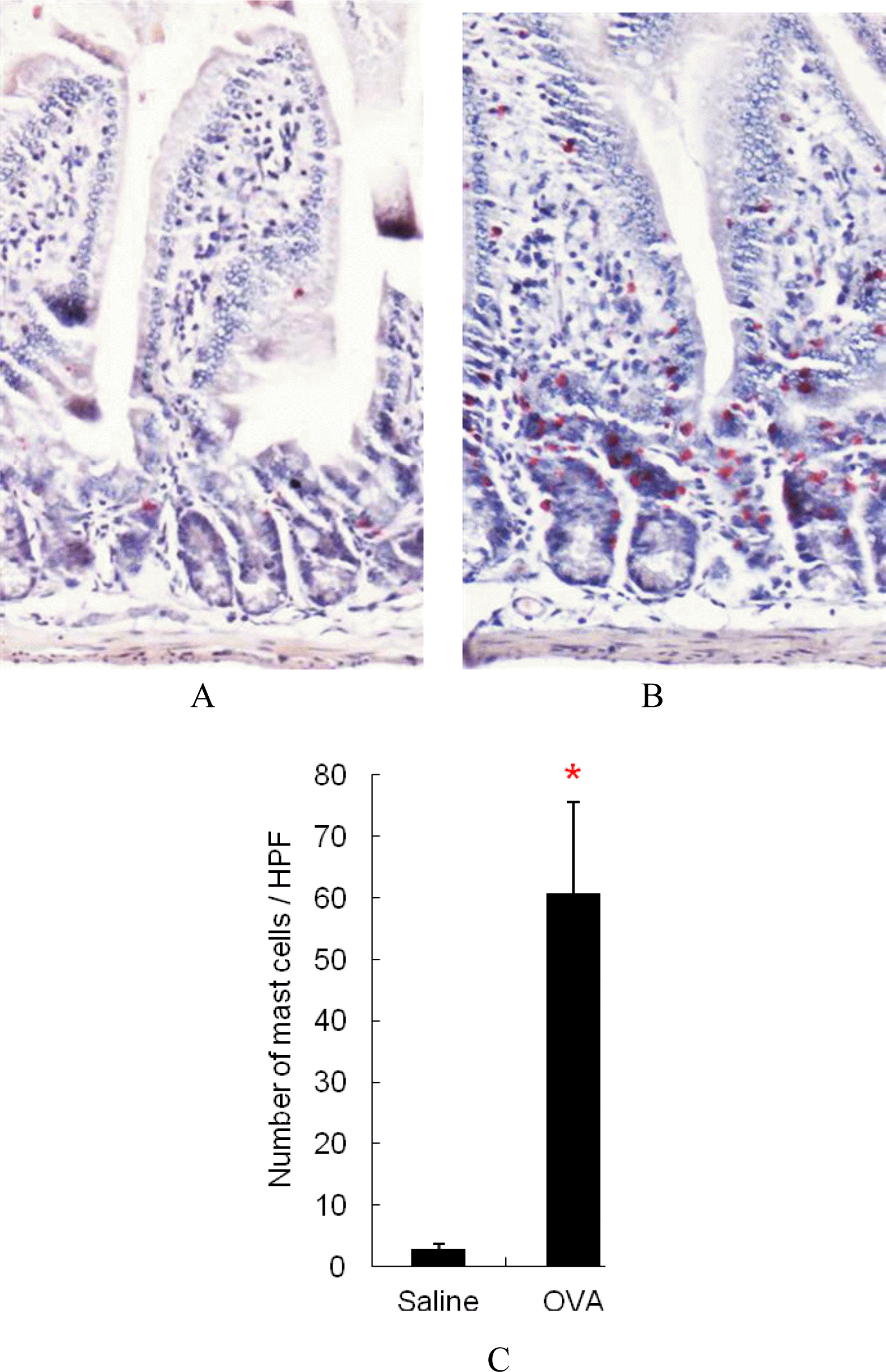
Figure 3 showed that the jejunum of OVA-sensitized mice contained highly increased levels of mast cells compared with sham-sensitized control mice and mast cells in jejunum of OVA-sensitized mice were predominantly localized to lamina propria region (Figure 3A and B). Furthermore, as shown in Figure 3C, mast cell levels in the jejunum of OVA-sensitized mice were significantly higher than that of sham-sensitized mice (p<0.05).
Analysis of bacteria diversity in colon samples using PCR-DGGE
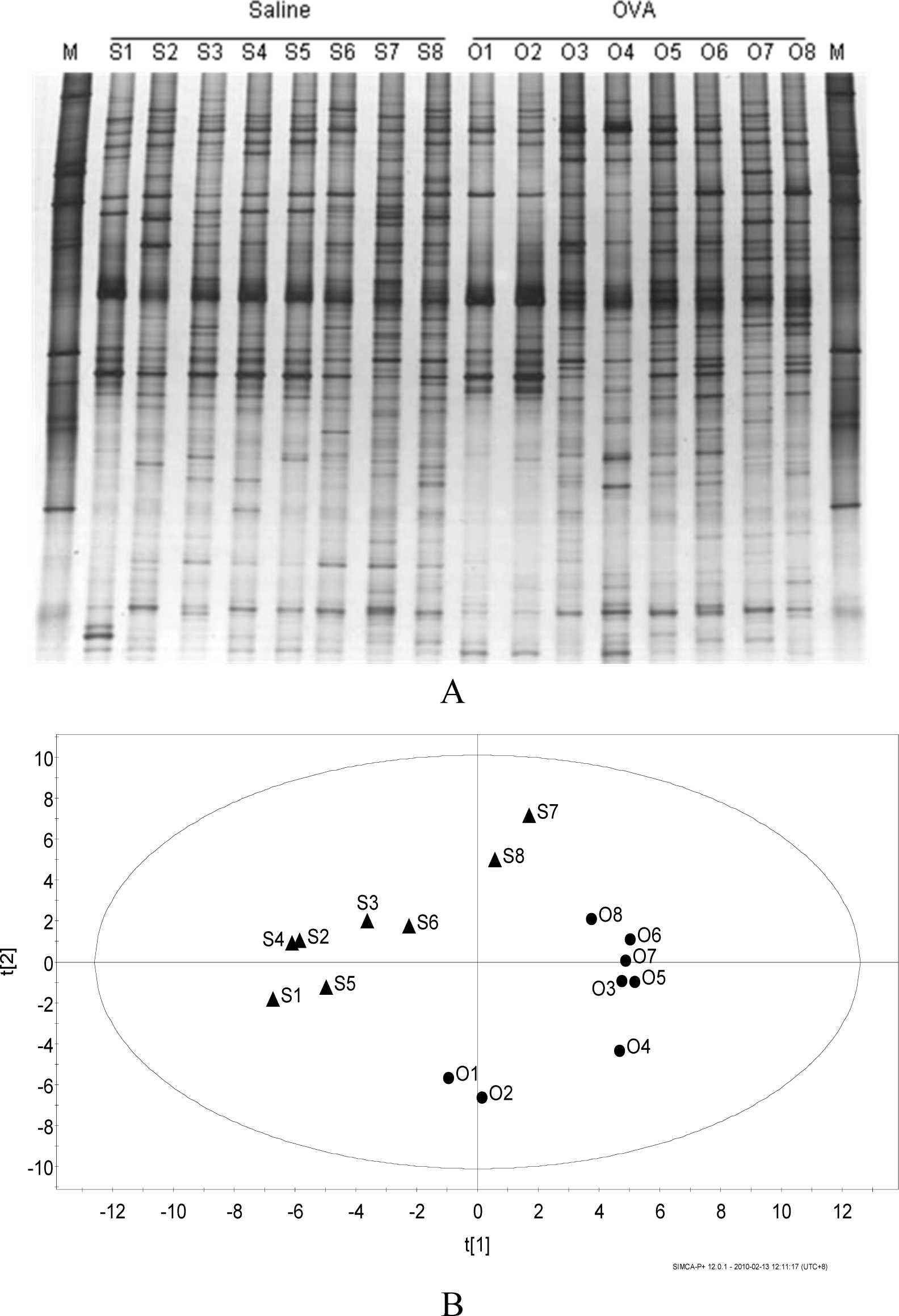
Figure 4A showed no specific band appeared or disappeared on the bacteria DGGE profiles of the content of colon. Principle component analysis for the DGGE profile (Figure 4B) showed that samples from group of OVA-sensitized mice and group of sham-sensitized mice were scattered into two different regions, indicating certain inter-treatment variation.
Quantitative real-time PCR analysis
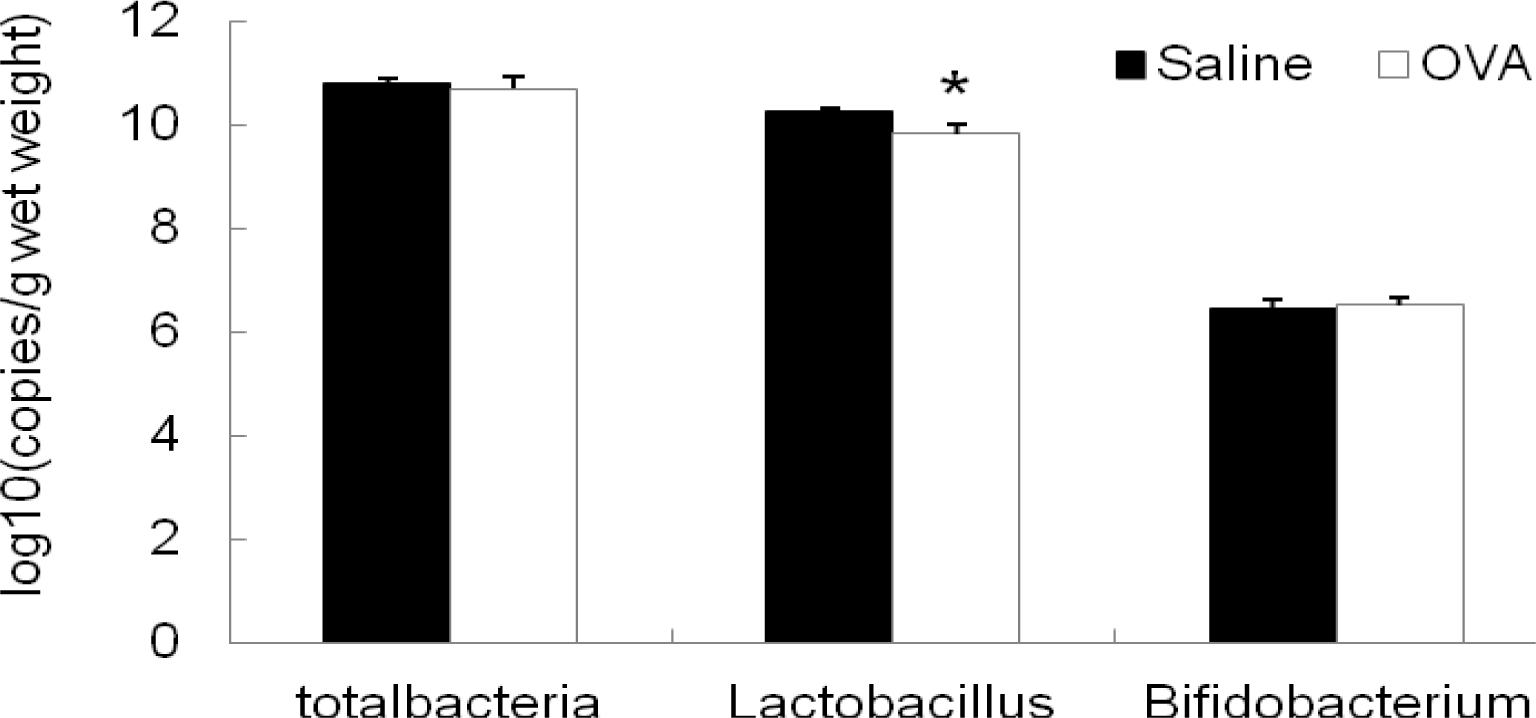
As shown in Figure 5, the concentration of Lactobacillus 16S rRNA gene copies was significantly lower in colon samples of mice with OVA-sensitization than those of sham-sensitized mice (p<0.05). However, no significant changes in the number of total bacteria and Bifidobacterium in the colonic content were found between OVA-sensitized mice and sham-sensitized mice.
DISCUSSION
This study presented a mouse model of OVA-induced allergic diarrhea according to Brant et al. (2003) under sub-barrier system. The hygiene hypothesis (Strachan et al., 1989) of allergic diseases indicates that strict environmental facilities, such as SPF condition, are required to induce Th2 responses. However, SPF conditions are expensive to establish and maintain. Therefore, a facility, which is in line with international standard and having low cost, instead of SPF condition is need to establish. Previous studies have showed that laminar flow cabinet in common laboratory animal facility can provide a local SPF condition and can be successfully used to breed nude mice (Li et al., 2003) and diabetes Scid mice (Li et al., 2005) which are both immune-deficient mice. In this study, BALB/c mice were maintained in the laminar flow cabinet and found to successfully develop diarrhea after repeat OVA challenges, although diarrhea occurred in this study was later and less sever. These data suggested that an acute allergic response could be induced in these mice under sub-barrier system.
We further demonstrated that diarrhea occurred in this study was allergic diarrhea. Food allergies are adverse immunological responses to food proteins. Allergens generally induce a shift in the Th1/Th2 balance toward Th2 immunity (Helm et al., 2003). In the Th2-biased cytokine microenvironment, IgE is produced by B cells and binds to its receptor on mast cells. Upon subsequent ingestion, antigenic fragments may interact directly with receptor-bound IgE on mast cells and trigger the release of the preformed and newly formed chemical mediators responsible for clinical symptoms (Prioult et al., 2005). Previously, Brant et al. (2003) observed that oral OVA-induced food allergic diarrhea is largely mast cell and IgE dependent. In this study, we also found that levels of OVA-specific IgG1 and total IgE in sera as well as the number of mast cells in jejunum greatly increased after repeat challenges. These data suggested that diarrhea developed in this study was allergic diarrhea.
Results of this study led to an important discovery that OVA-sensitized mice and sham-sensitized mice were discriminative for colonic microbial populations after the induction of diarrhea. Previous studies have showed that gut microbiota composition might be involved in allergic diseases. Wang et al. (2008) showed that there was a reduction in diversity of fecal microbiota in the early stage of infants with atopic eczema. In this study, no significant difference in diversity of colonic microbiota was found between healthy mice and diarrheal mice. However, principle component analysis of DGGE profile showed that samples from group of OVA-sensitized mice and group of sham-sensitized mice were scattered into two different regions, indicating certain inter-treatment variation.
Another finding of this investigation was that numbers of Lactobacillus in the colon of mice decreased as allergic diarrhea occurred. Previously, many studies have revealed that numbers of lactobacilli and bifidobacteria in the intestine of allergic disorders reduced as allergic reactions develop, while numbers of coliforms and Staphylococcus aureus increased (Bottcher et al., 2000; Bj├Črkst├®n et al., 2001). Supplementation of lactobacilli and bifidobacteria improved the balance of the intestinal microbiota, which was beneficial for the prevention of allergic diseases, including atopic dermatitis, asthma, allergic rhinitis and allergic diarrhea (Forsythe et al., 2007; Sunada et al., 2007; Schouten et al., 2009; Won et al., 2011). In this study, our results showed that copies of Lactobacillus 16S rRNA gene in colon samples of OVA-sensitized mice was greatly reduced, suggesting a close relationship between development of allergic diarrhea and reduction of Lactobacillus in the colon. At present, increasing evidences suggested that some cell components in probiotics such as immunostimulatory sequence oligodeoxynucleotides (ISS-ODNs) suppressed the Th2 immune responses (Takahashi et al., 2006; Kim et al., 2008) or inhibited the IgE production (Iliev et al., 2008; Hougee et al., 2010), resulting in the alleviation of allergic diseases. In addition, it was reported that mast cells express Toll-like receptor (TLR) (Bischoff et al., 2007), suggesting that gut microbes may interact directly with TLR on mast cells and regulate allergic responses (Bashir et al., 2004). However, further studies are required to uncover the mechanisms of the allergy-preventing effect of gut microbiota.
CONCLUSIONS
In conclusion, this study presented a mouse model of OVA-induced allergic diarrhea under sub-barrier system and showed that the number of colonic Lactobacillus reduced after induced diarrhea. These results suggested that food allergic diarrhea is associated with colonic Lactobacillus and that the reduction of colonic Lactobacillus numbers may be an important factor in the development of food allergic diseases. But further studies are warranted to evaluate the role of gut microbiota in the development of food allergic reactions.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







