





INTRODUCTION
In the transition period, dairy cows experience oxidative stress and are susceptible to metabolic disorders and immunologic challenges [1,2]. At the time of early lactation, cows go through a period of negative energy balance (NEB) which may lead to oxidative stress and health problems in these animals [3]. Early lactation NEB not only decreases milk yield but also increased concentrations of non-esterified fatty acids (NEFA) and beta-hydroxybutyrate (BHBA) [4,5]. NEFA in the blood is mobilized from adipose tissue while BHBA is an intermediate metabolite of fatty acid oxidation [6]. NEFA and BHBA are considered to be biomarkers of NEB, and oxidative stress during the transition period [7,8]. The body condition score (BCS) is an assessment of body fat and reflects the NEB of Holstein dairy cows [9,10]. It was reported that the BCS of cows were associated with differences in milk production and composition, reproduction, and health [7,11]. Cows with higher BCS at calving had higher concentrations of NEFA [12,13] and BHBA [13] in early lactation compared these with lower BCS. Cows with higher BCS are more sensitive to oxidative stress [14,15]. Oxidative stress is mainly derived from an imbalance between reactive oxygen metabolites and antioxidants and the concentration of some oxidative status indices such as malondialdehyde (MDA) was higher in cows with higher BCS before calving [15].
But the change of BCS during the transition period and their effects on the health and subsequent performance of dairy cows was few systematically studied. Therefore, the relations between the BCS and milk production, lipid metabolism, oxidative status of dairy cows were investigated in the present study. Consequently, this study may be used as reference for the dairy farms to improve the animals’ health status and performance by adjusting prepartum BCS.
MATERIALS AND METHODS
Animals and experimental design
The trial was carried out at a commercial dairy farm with about 5,000 lactating cows in Dongying City, Shandong Province, P.R. China. The average milk yield per lactation of the cows was more than 40 kg (305 day). The BCS (5-point scale) was individually recorded 21 days before calving, on the calving day and 21 days after calving [9]. Multiparous Holstein cows (second or third lactation) were selected according to their BCS at 21 days before calving and divided into 4 groups (28 animals per group): medium BCS (3.0 to 3.25, MBCS), high BCS (3.5 to 3.75, HBCS), higher BCS (4.0 to 4.25, HerBCS), and highest BCS (4.5 to 5.0, HestBCS). The cows were given access to total mixed ration and water ad libitum. The cows received a far-off diet (1.33 Mcal/kg) from −60 d to −22 d, and a close-up diet (1.50 Mcal/kg) was offered during the last 21 d. From 4 to 6 days postpartum, cows were supplemented with propylene glycol (PG, 300 mL) as a gluconeogenic precursor for both treatment and prevention of ketosis during transition. Cows were fed lactation diet I until 21 d (1.78 Mcal/kg) and lactation diet II (1.83 Mcal/kg) from 22 d to 100 d and milked four times at approximately 6:00, 12:00, 18:00, and 0:00; the individual milk yield was recorded every 3 days. The compositions of the diets are shown in Table 1 [16]. All of the animal experimental procedures were approved by the Institutional Animal Care and Use Committee at Jilin Agricultural University.
Blood and milk samples
Blood was sampled via the coccygeal vein after morning feeding at days 21, 14, and 7 prepartum (precalving), day 0 (calving), and days 7, 14, and 21 postpartum (after calving). Serum was collected in (1.5 mL) after centrifuging at 2,000×g at 4°C for 10 min and stored at −20°C until analytical determination. Colostrum was taken immediately after calving, and the levels of lactoprotein, lactose, solid-not-fat, and ash were analyzed by a milk analyzer (Foss Electric, Hillerød, Denmark) while the density and immune globulin concentration were analyzed by a refractometer (Yucheng To You Optical Instruments Co. Ltd., Shandong, China). The concentrations of NEFA, BHBA, superoxide dismutase (SOD), MDA, catalase (CAT), vitamin A (VA) and vitamin E (VE) in serum were analyzed using commercial bovine specific enzyme-linked immunosorbent assay kits (Shanghai Enzyme-linked Biotechnology Co. Ltd., Shanghai, China) according to the instructions. Reaction is terminated by sulphuric acid. The color change is measured spectrophotometrically at a wavelength of 450 nm (Multiskan FC, Thermo scientific, Waltham, MA, USA) and calculated by comparing the optical density of the samples to the standard curve, respectively.
Statistical analysis
All of the data are presented as the means±standard error and were subjected to one-way analysis of variance (colostrum composition) and general linear model by SPSS computer software (SPSS Inc., Chicago, IL, USA). The specify model included BCS (MBCS, HBCS, HerBCS, and HestBCS), Time (precalving, calving, postcalving) and their interactions (BCS× time) as fixed effects and cow within the treatment as random effect. Statistical significance is defined when p values are less than 0.05. Pearson correlation coefficients among the detected parameters were also calculated.
RESULTS
Colostrum composition, milk yield, and body condition score loss
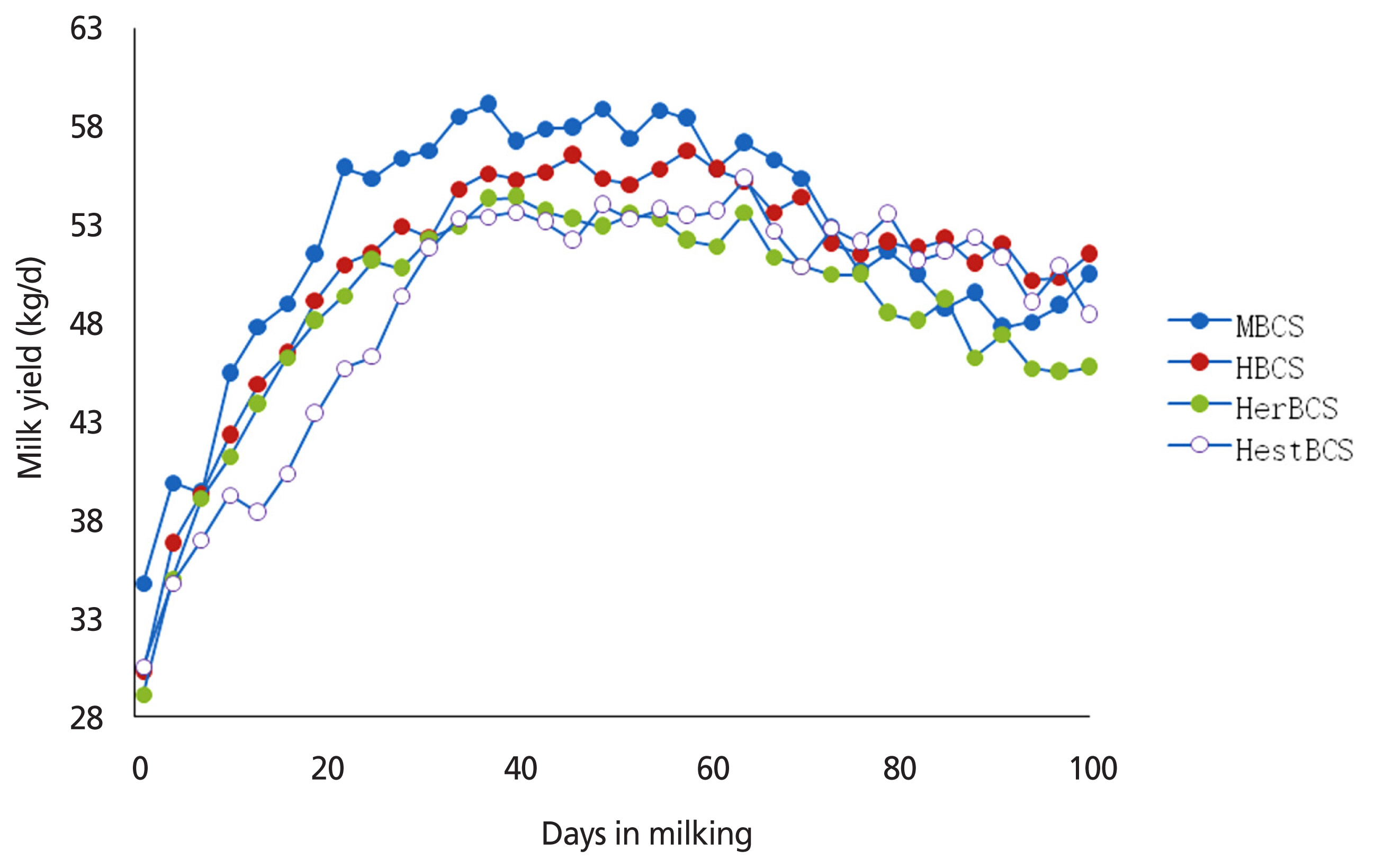
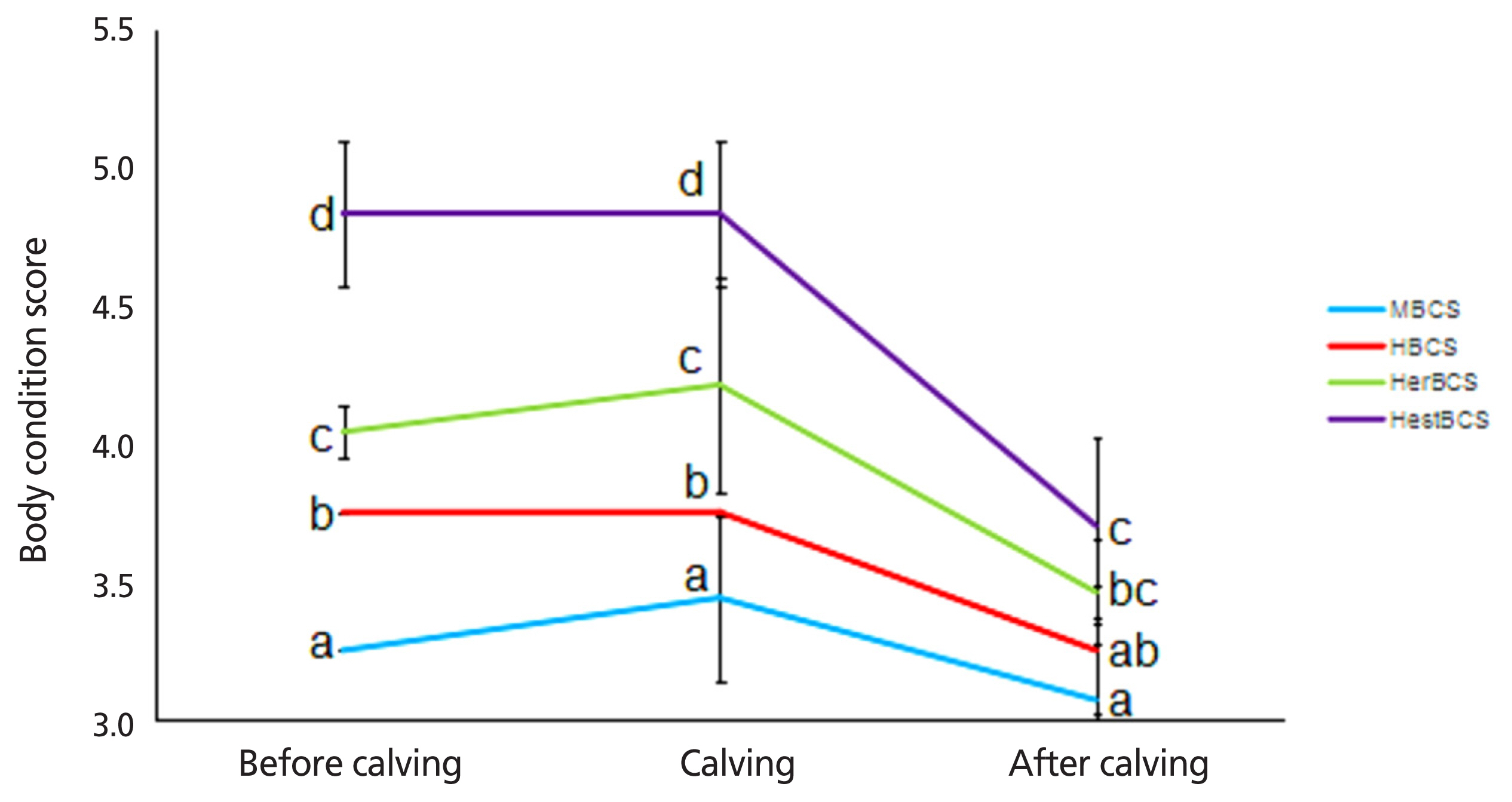
The density of colostrum and the levels of immune globulin and lactoprotein of cows in the HestBCS group were higher (p<0.05) than those in the other groups (Table 2). The milk yields of cows in the MBCS, HBCS, HerBCS, and HestBCS groups peaked (≥50 kg/d) at days 19 to 82, 22 to 100, 25 to 73, and 31 to 97, respectively. The average milk yields (0 to 100 days) were 52.35 kg/d, 50.94 kg/d, 48.59 kg/d, and 48.78 kg/d, respectively (Figure 1; Table 2). In addition, the present data showed that the animals in the HestBCS group had a greater BCS loss after calving (Figure 2). The percentages of lost BCS were 23.62% (HestBCS), 14.36% (HerBCS), 13.33% (HBCS), and 5.53% (MBCS) (Figure 2).
Lipid metabolism and oxidative status
NEFA was affected by both BCS (p<0.01) and time (p<0.01), while BHBA was only affected by BCS (p<0.01). There were no BCS×time effects on NEFA (p = 0.21) or BHBA (p = 0.97) during the transition period (Table 3). The serum levels of NEFA in cows in the HestBCS group on the calving day and after calving were all higher than those observed in other groups (p<0.05). In addition, the concentrations of BHBA were the highest (p<0.05) in HestBCS on precalving, calving, and postcalving days. Table 4 shows that MDA, SOD, and VA were affected by both BCS and time (p<0.05). There were BCS×time effects on SOD and VA during the transition period (p = 0.05). The concentrations of MDA, SOD, CAT, VA, and VE (except during precalving) in HestBCS (4.5 to 5.0) cows were all higher (p<0.05) than those in the other groups. Positive Pearson correlation coefficients were observed for NEFA (r = 0.260), BHBA (r = 0.496), MDA (r = 0.700), SOD (r = 0.606), CAT (r = 0.342), VA (r = 0.245), and VE (r = 0.448) to BCS (Table 5).
DISCUSSION
In early lactation, dairy cows experience a period of NEB due to the sudden increase in requirements of the mammary glands for milk production [17]. The mobilization of adipose tissue is associated with decreasing body weight (BW), and cows in transition with higher BCS lose more BW [18,19]. Similar results were observed in our study as the cows in the HestBCS group had the greatest BCS loss at 21 d after calving. Loker et al [20] suggested that lower production is associated with greater BCS. In our study, animals in the HestBCS group had a delayed peak milk yield. However, Dechow et al [21] found that fatter cows with more loss of BCS in early lactation have greater milk production. Milk composition is positively associated with BCS, and higher calving BCS means more protein and lactose during early lactation [22,23]. In our research, a higher level of lactoprotein was observed in HestBCS cows.
The NEFA and BHBA are commonly measured as indices of NEB and fat oxidization, and their concentrations during the early postpartum period are associated with BCS [10,19, 24,25]. The NEFA concentration in blood can be a reason for adipose tissue mobilization due to excess acetyl CoA generated from β-oxidation into ketone bodies, this leads to the loss of BCS [26,27]. In the present study, more rapid BCS loss and higher levels of NEFA and BHBA after calving were found in HestBCS cows, which is consistent with the results of Adrien et al [28]. Barletta et al [19] indicated that cows that lost BCS during the transition period had higher concentrations of NEFA and BHBA and more health events during the lactation. However, Mansouryar et al [23] fund that there was no significant differences in concentrations of NEFA and BHBA in high and normal BCS cows in early lactation. A BCS of 3.5 or higher at calving is associated with increased risk for ketosis [15], and a concentration of BHBA ≥1,200 μmol/L is considered as a sign of clinical ketosis [29,30]. However, no cows in the present study were diagnosed as ketotic at 1 week postpartum, as the BHBA concentrations were all lower than 1,200 μmol/L. This may be because choline (STA-CHOL) was added in the close-up diet [31] and PG was orally fed to cows at 4 days postpartum [32,33].
The MDA is the end product of the oxidative destruction of lipids, and its content can indicate the oxidative status of dairy cows [34,35]. In the present study, the highest level of MDA was found in HestBCS cows and MDA were increased on the calving day and decreased after calving. These data demonstrate that cows suffered the most serious oxidative stress on calving day and cows with higher BCS suffered much more serious oxidative stress than the other BCS cows before or after calving. It is probable that a negative association between BCS and dry matter intake in early lactation [36]. To maintain the requirements for milk production, prepartum cows with higher BCS experience more serious NEB [17].
The SOD is considered to be the first defense against pro-oxidants that convert the superoxide (O2−) to hydrogen peroxide (H2O2) [37] and Bernabucci et al [38] used SOD in blood to assess the oxidative status of transition dairy cows. In the current study, HestBCS cows had the highest SOD concentrations. And the concentration of SOD was commonly higher on calving day which was similar to the report of Bernabucci et al [15]. CAT is a critical antioxidant enzyme that prevents the accumulation of H2O2 and is considered to be an index of antioxidative status [39,40]. In the present study, CAT and MDA showed positive correlations, which is consistent with a previous study [35].
The VA and VE are fat-soluble membrane antioxidants that are important for enhancing antioxidant defense systems against oxidative stress [41,42]. Jin et al [43] reported that a diet supplemented with VA increases the concentration of serum SOD and CAT, which is consistent with the results of the current study. The VE supplementation reduces oxidative damage in the livers of heifers [44] and mitigates oxidative stress in crossbred cows [45]. In the present study, an oxidative (NEFA, BHBH, MDA)-antioxidative (SOD, CAT, VA, VE) balance was obviously observed in the cows, which might be the result of homeostatic control.
CONCLUSION
This study indicates that the milk yield, lipid metabolism, and oxidative status of Holstein cows are related to prepartum BCS and BCS loss during the transition period. Holstein cows with MBCS have better milk yield performance after calving whereas the animals with HestBCS suffer greater BCS loss and higher oxidative stress. Our data may provide some theoretical basis for improving the postpartum health and performance of dairy cows by adjusting prepartum BCS.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







