





INTRODUCTION
Corn grain is a main cattle energy source ingredient to produce commercial concentrate and total mixed ration (TMR). Thus, improving corn utilization is one of the fundamental factors to increase efficiency of cattle production. But the grinding level of the corn grain is used with limited information in terms of rumen fermentation patterns and blood metabolism [1].
In theory, grinding of corn increases the feed surface area and increases the digestibility of grain, resulting in improved grain utilization and production in monogastric animals [2]. The grinding method is also commonly utilized by the feed industry to produce concentrate feeds and TMR while preventing feed materials separation [3]. However, a very small particle size obtained fine grinding also results in certain disadvantages, e.g. stomach ulcer occurrence, significant amount of dust, which can reduce feed intake, etc. [3]. For the ruminant, several grain processing techniques including the grinding method have been examined to improve feed utilization [4,5]. Energy utilization in the rumen of beef cattle fed whole corn grain (WC) increased compared with those fed finely grinding corn grain (FC) [6]. Also, Reinhardt et al [7] reported a 7% feed efficiency increase when Holstein steers consumed WC as concentrate feed compared to FC feeding. However, there are difficulties in determining ruminal fermentation and physiological mechanism due to digestion by ruminal microbes, microbial synthesis and environmental factors including temperature, humidity, and other factors, etc., so further studies about changes in ruminal function as influenced by particle size of corn grain are still required [8].
Recently well-established experimental resources including at least no-stress conditions based on temperature and humidity levels have been provided for in vivo studies [9,10]. This may be well founded because a high level of temperature and/or humidity often increases the body temperature and energy requirements for maintenance resulting in unexpected metabolic data from in vivo studies using experimental animals [10].
Therefore, we examined the effects of different corn particle size on ruminal fermentation and blood metabolites of Holstein steers fed TMR as a basal TMR. The present in vivo study was performed under temperature and humidity-controlled conditions to minimize undetectable errors caused from the environmental factors.
MATERIALS AND METHODS
Animals and diets
The experimental protocol describing the management and care of animals was reviewed and approved by the Daegu University Institutional Animal Care and Use Committee (DUIACC-2016-001-0310-004). Four Holstein steers weighing an average 592 (standard error [SE]±29.9) kg and fitted with 100 mm i.d. ruminal cannula were used in a 4×4 Latin square experiment. The animals were then housed individually in 3×3.2 m pens in an auto temperature and humidity modulated chamber (24°C and 60% for 22 h/d, respectively) with water and a trace-mineralized salt block available throughout the experiments. The steers were weighed before each period. They were provided with two equal meals daily at 09:00 and 17:00 h.
The steers were offered TMR as a basal diet ad libitum during the adaptation period, and then the DM intake was restricted to 90% of the ad libitum intake (mean 1.66% of body weight). A basal diet (% dry matter [DM]) consisted of commercial corn based concentrate (48.5), barley brewer’s grain (25.5), rice straw silage (13.4), perilla meal (6.7), corn flake (2.8), soybean curd process (2.8) and limestone (0.3). The 20% of energy intake from the basal diet was replaced by one of the grinding levelled grains of Kwangpyeongok, Korean domestic corn (Zea mays), as follows: no replacement of corn (i.e. TMR only; control), WC, coarsely ground corn grain (CC), or FC, respectively. The treatments used same corn as whole corn grain, coarsely ground corn grain and finely ground corn grain of Kwangpyeongok, and replaced by 20% of energy intake from the TMR. Therefore, all treatments had the same in the chemical composition. Processed corn grain samples were prepared as follows: coarsely ground corn grain was prepared by grinding through 6.0 mm and then 1.3 mm mesh screens of a Wiley mill (Model 4; Thomas scientific, Swedesboro, NJ, USA), i.e. the coarsely ground corn grain was collected between 1.3 mm and 6.0 mm mesh screens. Finely ground corn grain was prepared by grinding through 3.5 mm mesh screen of a Wiley mill, because this is a commercially available particle size when compounding a TMR in a Korean feed company. Feed DM intake was changed according to the body weight of the steers determined at the beginning of each period.
Sampling procedures and chemical analysis
Each period lasted 21 days and feed intake was recorded daily. Representative samples of feeds were collected on days 15 to 21, pooled, subsampled, dried, and ground through a 2-mm screen in a Wiley mill (Model 4; Thomas scientific, USA), and analysed for moisture, crude protein, ether extract, and ash according to the procedure of AOAC [11]. The concentration of neutral detergent fiber corrected for residual ash was determined with heat-stable amylase and sodium sulphate according to the method of Van Soest et al [12], while the concentration of acid detergent fiber corrected for residual ash was determined according to the procedure of AOAC [11]. The chemical composition of experimental feeds is shown in Table 1.
Rumen fermentation characteristics were assessed by collecting ruminal samples just before the morning feeding (0 h) and at 1, 2, 3, 4, 5, 6, 7, and 8 h post-feeding on day 21. Immediately after collection, the samples were filtered through eight-layer cheesecloth and then ruminal pH was measured. For volatile fatty acid (VFA) determination, 1.0 mL of saturated HgCl2 solution and 4.0 mL of 1 M NaOH solution were added to 10.0 mL of rumen fluid. For ammonia-N determination 0.3 mL of 50% H2SO4 was added to 15.0 mL of rumen fluid. Samples were stored frozen at −20°C until analysed. Ammonia-N and VFA were determined according to Choi and Oh [13].
Blood samples were taken by venipuncture from the jugular vein at the end of the experiment periods prior to afternoon feeding (17:00 h). Blood was collected into 10-mL vacuum tubes (BD Vacutainer, Becton & Dickinson, Franklin Lakes, NJ, USA) without anticoagulant. The samples were allowed to clot for 24 h at 4°C and centrifuged (2,000×g for 15 min at 4°C), and serum was stored at −70°C until further analysis. Blood samples were analyzed by an automatic blood analyzer (Modular Analytics, Roche, Wiesbaden, Germany). These included total protein, glucose, blood urea nitrogen (BUN), albumin, creatinine and triglycerides.
Rectal temperature was determined using a portable multichannel thermometer recorder (Thermo Recorder, TR-71U, T&D Corporation, Matsumoto, Japan) just before the morning feeding on day 21.
Statistical analysis
Data for steers within each treatment were averaged and analysed using the general linear model procedure of SAS (SAS Inst. Inc., Cary, NC, USA) with steer as random effects, and treatments as fixed effects. Duncan’s multiple range test was used to interpret any significant differences among the mean values of the treatments. Differences among treatment groups were considered significant if p<0.05.
RESULTS AND DISCUSSION
Feed intake and animal condition
Grinding method of corn grain did not increase (p>0.05) TMR and total feed DM intake (Table 2). On an average, the experimental animals consumed 8.7 (SE±1.61) and 10.3 (SE±1.87) kg of TMR and total feed DM, respectively. In the present study, feed DM intake was restrictively fed based on the body weight of each period. Also, we designed the trial so that the animal consumed all the feed with no ort for accurate interpretation of metabolic data including rumen fermentation and blood metabolites. Thus, the present intake data provided limited information. The substitution of corn grain and the grain grinding method did not change body temperature (mean 38.2°C; Figure 1) because the experimental animals in the present study were housed under no stress conditions based on temperature and humidity index (THI = 71.5) according to the suggestion by Bilby et al [14]. Previously, severe heat stress (THI = 81.4; 30.0°C of temperature and 70% of humidity) numerically increased the body temperature and the energy requirement for maintenance of cows [9] while decreasing the DM intake [15]. Coppock [16] reported that increases in environmental temperature led to heat stress to ruminants, resulting in high body temperature and respiration rate. Also, high body temperature caused a decrease in body weight [17] and was an important parameter for investigation of reproduction, milking and growth [18]. Thus, well-controlled experimental resources including at least no-stress conditions based on temperature and humidity should be provided for accurate interpretation of nutritional and physiological data from in vivo animal studies.
Rumen fermentation
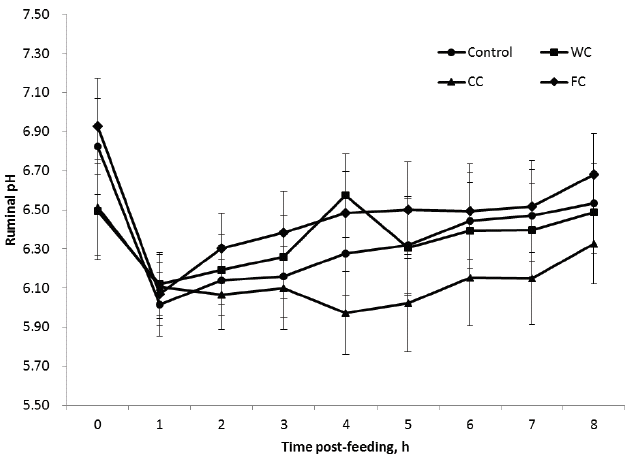
Different corn particle size significantly (p<0.05) affected the mean ruminal pH and individual VFA, whereas such grinding did not change mean ammonia N and total VFA concentrations (p>0.05) (Table 3). Ruminal pH for CC was lower than the other groups, in particular at 2 h post-feeding and thereafter (Figure 2). At 1 h after feeding ruminal pH for FC was numerically the lowest but increased to be the highest during the rest of the feeding cycle compared with the other groups. This is consistent with Lee et al [1] in which in vitro ruminal pH from incubation with ground corn grain was lower than that with whole corn grain. However, compared with the present in vivo study, they also showed inconsistent results, i.e. similar pH changes in incubation with coarsely- and finely-ground corn grains occurred [1]. It was likely that the present pH results obtained some unexplained experimental errors because reduction of particle size by grinding was generally related to decreased ruminal pH [1,19]. But, the different results between the studies may be a result of differences in the actual particle size of ‘coarsely-’ or ‘finely-’ ground corns in the previous study [1]. In the study of Lee et al [1], coarsely- and finely-ground corn grains were prepared by grinding through 4 mm and 1 mm mesh screens, respectively. It seemed that the ‘coarsely’ ground corn from the 4 mm mesh filtration contained a considerable amount of smaller than 1 mm ‘coarsely’ ground corn grain [1]. Our presumption may be supported by the observation of little differences in digestibility between finely (<2.5 mm) and coarsely ground (3.5 mm) corn grains when the particle size of corn grains are similar [20]. In the present study, to avoid such experimental error, we used the ground corn grains from a size between 1.3 to 6.0 mm for the coarsely ground corn grains and a size of smaller than 3.5 mm for the finely ground corn grains, respectively, even though some particle sizes may still have coincided (for details, see Materials and Methods). The reason for relatively high pH value of FC was not clear, but may have resulted from too small particle sizes of FC, i.e. it may had insufficient retention time for microbial attack in the rumen. Thus, the present pH results imply that the grinding reduces the size of particles and the coarsely grinding processing increases the corn grain surface area for the microbial attack [21], expected to lead high starch digestion in the rumen [19]. Starch is supposed to be readily degraded by rumen microbes and therefore an expected higher pH in FC.
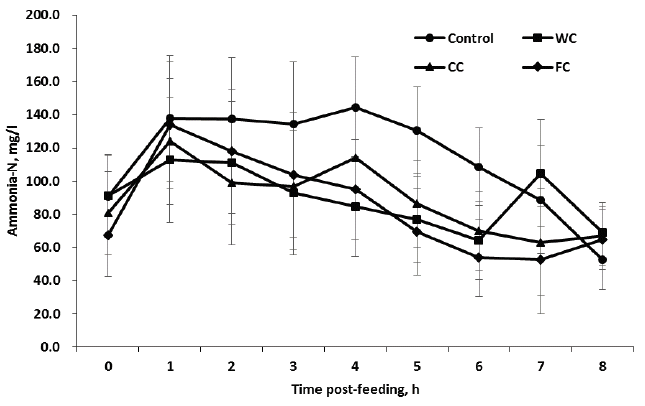
Feeding corn grains significantly (p<0.05) decreased mean ammonia N concentration compared to the control, but no changes in ammonia N appeared among corn grains fed groups (p>0.05) (Table 3, Figure 3). In the present study, N intake for control (1.60 kg/d, data not shown) was similar to those for WC (1.59) and numerically lower than CC (1.68) and FC (1.79), respectively. This should have resulted in lower ammonia N for control than the corn feeding groups since high N content available for microbial attack increases ammonia N in the rumen [22,23]. In the present study, high ammonia N in the control may have presumably occurred for two reasons, i.e. slower meal time of TMR and soluble N content in TMR. In general, ruminant animals have a better appetite for concentrate rather than forage including TMR [24]. Also, high soluble N in feeds is closely related to high concentration of ruminal ammonia N [25]. The previous observations were correct because the present diurnal pattern in ammonia N for control was similar till 1 to 2 h post-feeding and higher thereafter compared with the other groups despite lack of significance.
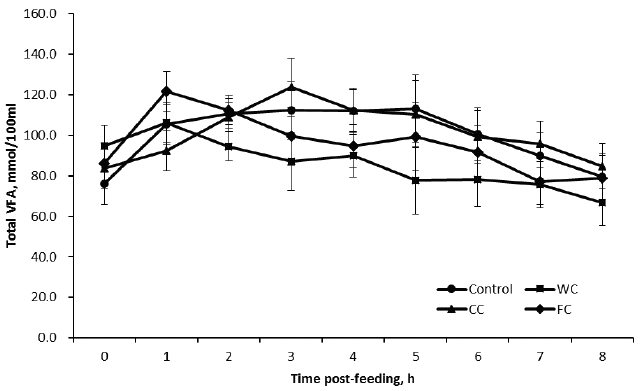
Feeding different particle size corn grains did not affect the total VFA concentration in the present study (Table 3, Figure 4). This is in contrast with the previous in vitro study by Lee et al [1] where coarsely and finely ground corn feeding were significantly higher than whole corn feeding. The discrepancy in total VFA between the present and previous studies may result from the differing experimental types as well as the particle size discussed above. An in vitro study is often likely to produce clear results because there are no animals involved and hence there is no chewing and ruminating steps, and no absorption of VFA [13]. Our observation agrees with the in vivo study of Reinhardt et al [7] where there was no change due to corn processing on ruminal fermentation as affected by total VFA concentration. The lowest acetate and the highest propionate were observed for CC, while the ratio of acetate to propionate was lower for CC than WC and FC. This is consistent with the previous results where corn grain grinding decreased the ratio of acetate to propionate in cattle [1]. Also, the ratio of VFA may be good for animal performance, in particular feed digestibility in the rumen because coarse grinding of corn grain (e.g. a grinding size of 3.2 to 7.9 mm) increased starch digestion [19] and DM digestibility [4]. Overall, in addition to ruminal pH data in the present study, the individual VFA and the ratio of acetate to propionate clearly supports our observation that coarse ground corn grain feeding may increase feed digestion in the rumen.
Blood metabolites
Feeding different particle size corn grains did not affect (p>0.05) blood metabolites except for triglyceride appearing to be higher for FC than the other groups (Table 4). Although blood albumin was not difference between treatments, blood albumin concentration was numerically higher for WC and CC (3.80 and 3.85 g/dL) than FC and control (3.73 and 3.68 g/ dL). However, this is not clear because, in general, blood albumin is an index of protein synthesis in a body, i.e. low albumin indicates relatively high protein synthesis [26]. However, the mean concentration of blood creatinine, the index of total muscle mass in a body [26], appeared to be rather similar to the patterns of albumin concentration in the treatments in the present study. Blood glucose was not different between treatments but numerically higher for FC than those for the other group. This is clearly supported by the high ratio of acetate to propionate for FC compared with control and CC in the present study in spite of low propionate concentration for FC (Table 3) because blood glucose concentration is closely related to the ratio of acetate to propionate in the rumen and related to glucose absorption in the small intestine [27]. Typically the carbon sources of fatty acids in ruminants is acetate rather than glucose [28]. However, glucose is definitely an important factor because Vernon [29] observed that, in addition to acetate, glycerol-3 phosphate is also required for triglyceride synthesis, which completely derives from glucose. Thus, the present higher triglyceride results for FC than the other groups were rather logical. Despite lack of significance, BUN is known to be increased by increasing absorption of ruminal ammonia N from the rumen wall [13]. The concentrations of ruminal ammonia N were, however, vice versa (Table 3). This may be not surprising because soluble N including ammonia N can escape from the rumen [30], and thus levels of ruminal ammonia N concentration and ruminal ammonia N absorption could be different [26].

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







