




INTRODUCTION
Feeding has been reported to induce reduction in the concentration of growth hormone (GH) in plasma [1,2], and cause growth hormone-releasing hormone (GHRH)-induced increase in GH in ruminant animals [1,3]. We hypothesized that post-prandial decrease in GH levels may be caused by increase in the levels of plasma short-chain fatty acids (SCFA), resulting from increased rumen fermentation and subsequent absorption of SCFA in the blood. This hypothesis was partly supported by reports showing that infusion of SCFA into blood, and/or an increase of SCFA in the medium bathing the anterior pituitary gland cells, suppressed GHRH-induced GH release [4–6]. However, it was also reported that an increase in mesenteric SCFA concentrations, which was mimicked by the infusion of mixed SCFA up to the post-prandial level, was not enough to suppress GHRH-induced GH increase [7]. From these results, it appears that physiological increase in plasma SCFA levels after feeding might not be enough to suppress the GH levels, highlighting the need for another explanation for these phenomena.
It is known that feeding decreases plasma GH, but increases plasma insulin levels, which indicates a reciprocal relationship between these two major metabolic hormones in adult ruminant animals [2]. In the adults of some animal species, a relationship between plasma GH and insulin concentrations has been reported [8–13]. Therefore, the post-prandial increase in plasma insulin concentration, which is usually accompanied by an increase in glucose concentration, might inhibit GH release from the anterior pituitary gland of the ruminant.
In the present study, we examined an alternative hypothesis for the post-prandial reduction in plasma GH levels: whether increase in plasma insulin levels is involved in GH reduction mechanisms. To accomplish this objective, we performed two experiments: intravenous infusion of insulin or glucose to increase the plasma insulin levels in goats.
MATERIALS AND METHODS
Animals
Male Shiba goats were treated in accordance with the “Guiding Principles for the Care and Use of Animals in the Field of Physiological Sciences”, as recommended by The Physiological Society of Japan. The experimental procedures were approved by the Animal Care Committee of Tohoku University (Approval number: 2010AgA-24).
The animals were fed alfalfa hay cubes (1.3%/total body weight [BW]) at 1000 h in the morning and were offered water and mineral salts ad libitum for a week before the blood sampling day. The nutrient composition of Alfalfa hay cubes (dry matter 87.5%) was crude protein 16.9%, ether extract 1.5%, acid detergent fiber 30.5%, neutral detergent fiber 39.1%. Alfalfa hay cubes were removed 16 hours before the blood sampling.
Intravenous insulin and glucose infusion, and blood sampling
Five male goats (23.28±3.52 kg BW) were used in this experiment. A polyethylene catheter was inserted into the left jugular vein of the animals 2 h prior to blood sampling for stabilization and reducing stress as our previous reports [14,15]. No acute increment of non-esterified fatty acid (NEFA) and cortisol before blood sampling indicated that 2 h is enough for stabilization in goats (Figures 1, 2). The catheter was kept in sterile 3.5% tri-sodium citrate. The blood was sampled from 0900 to 1300 h. Approximately 5 mL of blood was collected at each sampling time, 30 or 60 min before and 120 min after bolus injection of insulin (0.2 IU/kg BW), glucose (1.0 mmol/kg BW), or saline into venous blood through the catheter. All blood samples were kept in an ice bath until centrifugation at 3,000×g for 10 min at 4°C. The separated plasma samples were stored at −20°C until the analysis.
Hormone and metabolite assays
The concentrations of GH and insulin in the plasma were determined by radioimmunoassay, as described previously [16–18]. Cortisol concentration in plasma was analyzed by Cortisol ELISA kit (Enzo Life Sciences, Inc., Farmingdale, NY, USA). The plasma glucose and NEFA concentrations were measured using a commercial kit (Glucose C-II Test-Wako, NEFA C-Test-Wako; Wako Pure Chemical Co., Tokyo, Japan).
RESULTS
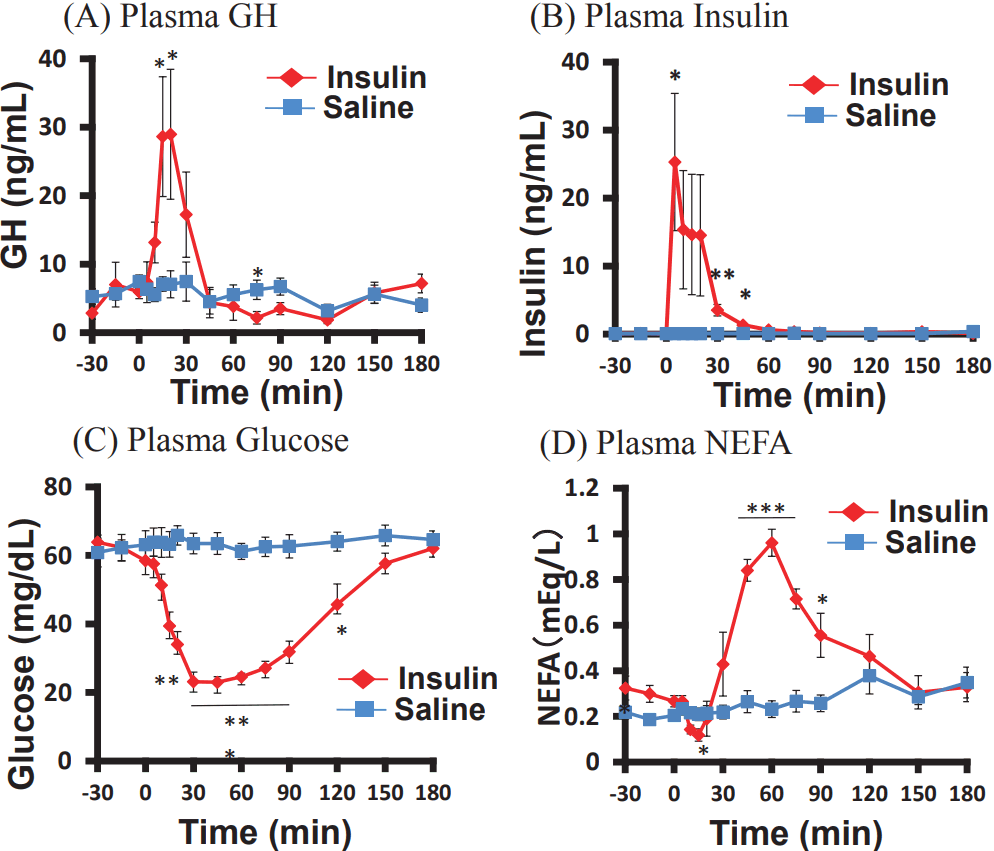
Effects of insulin injection on plasma GH, insulin, glucose, and NEFA concentrations
Compared to the control injection, the insulin injection, which significantly increased the plasma GH concentrations after 15 and 20 min (Figure 1A), significantly increased the plasma insulin concentrations 5 min and 30 to 60 min after the injection (Figure 1B). The blood glucose concentrations were significantly lower after 20 to 120 min (Figure 1C), whereas the concentrations of NEFA, 45 to 90 min after the injection were significantly higher, than the concentrations observed in the control (Figure 1D).
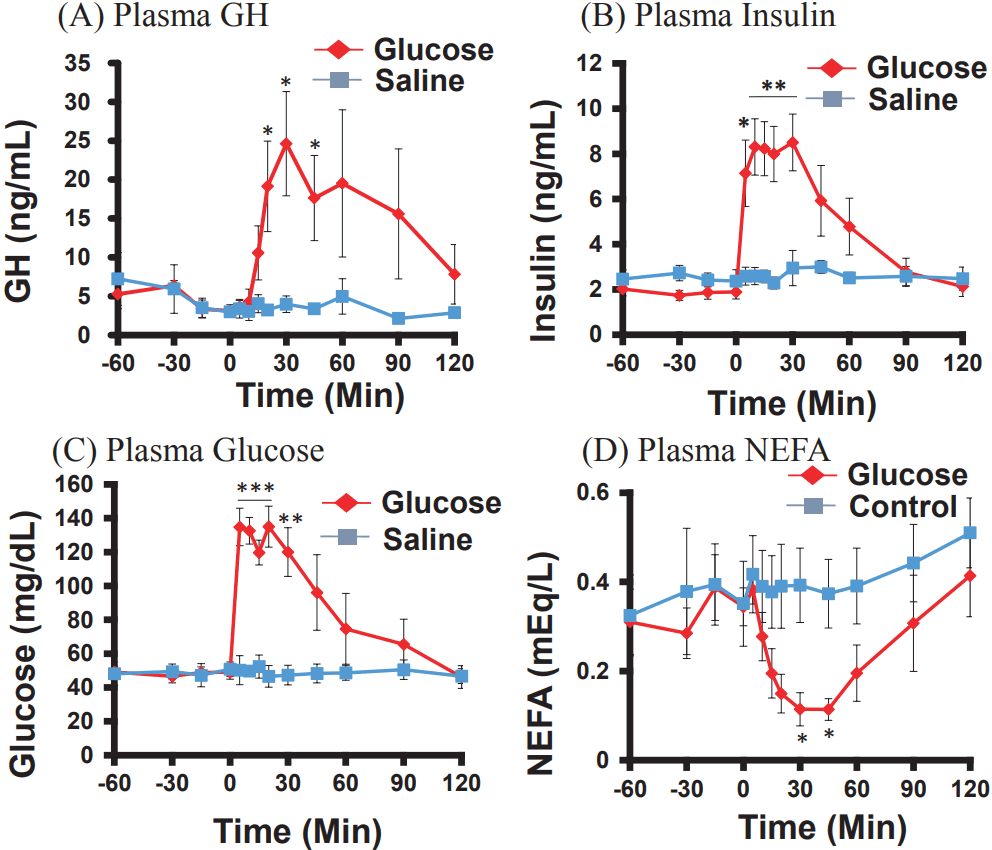
Effects of glucose injection on plasma GH, insulin, glucose, and NEFA concentrations
The concentrations of plasma GH were significantly higher than those in the control, 20 to 45 min after the glucose injection (Figure 2A). The plasma insulin concentrations were also significantly higher, 5 to 30 min after the injection, than in the control (Figure 2B). The blood glucose concentrations were immediately increased at 5 to 30 min (Figure 2C), whereas the plasma NEFA concentrations decreased, gradually but significantly, 30 and 45 min after the injection, as compared to the decrease in the control (Figure 2D).
DISCUSSION
The present study demonstrated that venous injection of insulin or glucose immediately and significantly increased the plasma concentrations of GH. This fact indicates that increase in plasma insulin concentrations always causes a rise in plasma GH concentrations, and that plasma insulin or glucose levels are not related to the post-prandial reduction in GH levels. The doses of insulin and glucose injection used in the present study have been employed in previous studies [19,20].
Volatile fatty acids increased after feeding have the inhibitory effects on GH secretion, and stimulatory effects on insulin secretion in sheep [2,6,7]. However, the significant increase in GH concentrations after the insulin infusion might have been caused by the stress from hypoglycemia and consequent stimulation of the hypothalamus-pituitary-adrenal (HPA) axis, because plasma cortisol concentrations were significantly increased by the insulin injection (Figure 3A). We previously reported that excitation of the HPA axis caused an increase in plasma GH concentrations [15,17]. Apelin and arginine vasopressin (AVP), the key stress hormones in ruminants, were reported to stimulate the secretion of adrenocorticotropic hormone [21–23], and subsequently cortisol was also demonstrated to stimulate GH secretion [14,17]. Lipopolysaccharide injection stimulated GH secretion in rats [24] and corticotropin-releasing hormone injection had the same effect in humans [25]. There are the possibilities on apelin and AVP secretions by insulin injection in the present experiment. All these findings suggest that stimulation of the HPA axis induced by insulin injection might be involved in the increased GH secretion. Further studies are needed to clearly demonstrate the action of insulin on GH secretion in hypothalamus and pituitary gland.
It also appears that insulin-induced hypoglycemia was involved in the elevation of GH concentrations. Insulin-induced hypoglycemia causes an increase in GH concentrations in humans [26]. It was also reported that restricted-feeding increased GH levels in ewes [27], suggesting that low-glucose conditions might have raised the plasma GH concentrations in the present study, although such a physiological phenomenon was not observed in sheep [28]. As GH acts to promote the release of glucose from liver through activation of the Janus kinase 2/Signal transducer and activator of transcription 5 (JAK2/STAT5) pathway in mice [29], it is possible that GH release was stimulated by insulin-induced hypoglycemia to aid the recovery of the lowered blood glucose levels.
The mechanism for increased GH concentrations induced by the glucose injection might be different from the mechanism of increase induced by the insulin injection, because plasma cortisol concentrations were not changed by the glucose injection. It was reported that glucose infusion induced an increase in plasma GH concentrations in dairy cows [30,31] and glucose stimulated the release of GH in goat anterior pituitary cells in vitro [32,33].
The NEFA was decreased immediately after the insulin injection, but it rapidly increased thereafter. This change was reciprocally accompanied by that of glucose concentrations (Figures 1, 2). Insulin promotes lipogenesis by increasing NEFA uptake by the adipose tissues [34]; however, GH has the opposite action [35]. It also appears that changes in plasma NEFA, as well as in glucose, might be involved in changing GH concentrations, because it has been reported that fatty acids directly inhibit GH secretion from cultured anterior pituitary cells [4].
In conclusion, GH secretion was significantly elevated, and not reduced, by the bolus venous injection of insulin as well as of glucose. Insulin, not glucose, also stimulated an increase in cortisol concentrations, indicating that increased GH secretion might be caused by stimulation of the HPA axis. Factors other than insulin might be involved in the post-prandial GH reduction in goats.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







