



INTRODUCTION
Rice straw is an important roughage resource for ruminants in many rice-producing countries. In China, the annual yield of rice straw is about 188 million tons (Guo et al., 2002). However, the utilization efficiency of rice straw is limited due to its low protein and energy value, high silica and consequently low digestibility. In many areas of China, rice straw is often burned in the field by farmers during harvest time, which occasionally induced serious environmental pollution. Therefore, in order to improve the utilization efficiency, many physical, chemical or biological pretreatments have been employed to improve rice straw nutritive quality, such as ammonification (Liu et al., 2002; Selim et al., 2004), urea treatment (Rahal et al., 1997; Vadiveloo and Fadel, 2009), steam spray (Liu and Ørskov, 2000; Weimer et al., 2003) and white rot fungi fermentation (Karunanandaa et al., 1995). Although these pretreatments showed some benefits for fermentation and animal production, additional costs and labor were required.
Previous studies have revealed that there was considerable genetic variation among varieties relative to the nutritive value of rice straw (Abou-El-Enin et al., 1999). A hybrid rice crossed with two low or high straw nutritive quality parents yielded higher dry matter digestiability (Singh and Singh, 1995; Sohane and Singh, 2000), which indicated that genetic improvement was an alternative approach to increase the nutritive value of rice straw. However, in most rice-producing countries the feeding value of rice straw was not considered in rice breeding programs. Mutagenesis is an important method for crop breeding, but few studies focused on the effect of gene mutagenesis on the nutritive value of rice straw.
A brittle mutant (BM) was obtained by means of 60Co-γ radiation of a japonica rice variety Zhonghua-11 (Wild type, WT). This mutant displayed normal phenotype similar to its wild type except for the fragility of its whole plant body. The gene fine mapping and sequence analysis have revealed that this mutational phenotype was caused by the loss-of-function mutation in OsCesA4, which encoded cellulose synthase catalytic subunit (CesA) (Yan et al., 2007).
Many physical and chemical factors, including particle size, cell wall structure, cell wall components and so on, could affect forage intake and digestibility in ruminants (Jung and Allen, 1995). Therefore, these mutative traits of BM rice straw may be favorable for its efficient utilization by ruminants. Hence, the botanical fractions, chemical compositions, stem histological structure and dry matter digestibility were investigated in both BM and WT in order to estimate whether a cellulose synthase gene mutation could improve the nutritive value of rice straw.
MATERIALS AND METHODS
The plant materials
BM and WT were sown on the same day at the experimental farm of Yangzhou University in the natural growing season from June to September. BM exhibited the fragile culm and leaf (Figure 1). Apart from the brittle phenotype, there were no differences between BM and WT in terms of other agronomic characters, including plant height, heading date, tilling ability and seed set. At the yellow maturity stage, which corresponded to 122 days of growth after sowing, both strains were hand-harvested 5 cm above the ground, subsequently the grains were removed. The two strains were collected in triplicate from three replicate plots.
Morphological proportion
During harvest stage, five plant samples at each experimental plot were collected, then these straws were divided into stem, leaf blade and leaf sheath fractions, respectively. Leaves were removed from stem internodes, then leaf blade and leaf sheath were separated. Nodes and internodes were pooled to constitute the stem fraction. All fractions were dried at 65°C for 72 h. The morphological proportion was estimated by weight of each fraction (on dry matter basis).
Chemical composition analysis
BM and WT rice straw were ground to pass 1 mm sieve for subsequent chemical analysis. Dry matter (DM), organic matter (OM) and crude protein (CP) were assayed according to AOAC (1999). Cellulose, hemicellulose, acid detergent lignin (ADL), acid insoluble ash (AIA), neutral detergent fiber (NDFom) and acid detergent fiber (ADFom) were measured following the procedures of Van Soest et al. (1991). NDFom and ADFom were calculated by subtracting the residual ash. Sulfite and heat stable amylase were not included in the procedure, and lignin content was determined using the sulfuric acid method. Hemicellulose was measured by subtracting ADFom from NDFom.
In situ digestibility
Three Holstein dairy cows with permanent rumen cannula fed alfalfa hay and corn silage based total mixed ration were used for the in situ digestion study. Nylon bags (7×14 cm; pore size, 45 μm) containing approximately 3.0 g ground rice straw were placed into the rumen immediately before feeding at 08:00 am. Bags were removed from the rumen after 0, 6, 12, 24, 48 and 72 h incubation for the determination of DM and rice straw components disappearance. These results were fitted to the exponential equation P = a+b(1-e−ct), where P represents the disappearance rate at time t, whereas a, b and c are constants. a means The readily digested fraction represented by a, the slowly digested fraction represented by b, the rate of digestion of the slowly digested fraction represented by c, and the potentially digestible fraction represented by a+b (Ørskov and McDonald, 1979). Effective digestibility (ED) was calculated by the equation ED = a+(bc/(c+k)), where k represented passage rate (McDonald, 1981). In this study, the hypothetical k value was 0.025. If the a value was negative, as noted by Wilman et al. (1996), it indicated a lag time before rapid degradation began, the length of the lag time was estimated as (1/c)ln[b/(a + b)] (McDonald, 1981).
SEM and TEM investigation
BM and WT stem samples (approximately 2 mm×5 mm) for scanning electron microscope (SEM) and transmission electron microscope (TEM) observation were taken from the second internode below the panicle. These samples were excised with a razor and immediately stored in 2.5% glutaraldehyde for approximately one month before post-fixation. After several rinses, the material was dehydrated in an ethanol series, critical point dried in CO2, spurted with gold and examined with SEM (Philips XL-30E ESEM, Holland).
Samples for TEM were rinsed, dehydrated as above and embedded in low viscosity resin. Thin sections (60 to 100 nm) were cut with a diamond knife, collected on copper grids and stained with 1% aqueous uranylacetate for 15 min followed by 15 min in Reynold’s lead citrate. The sections were examined with TEM (Phillips Tecnai 12, Holland).
Statistical analysis
Statistical analysis was performed using statistical program for the social sciences (SPSS) software for Windows (version 15.0). The difference in DM, chemical compositions, in situ DM digestibility and digestion constants between BM and WT were identified by independent-sample t-test. The constant a, b and c were evaluated by nonlinear regression model.
RESULTS
Morphological proportion
Morphological fractions of BM and WT rice straw are shown in Figure 2. The stem proportion of both strains was similar, accounting for 30% approximately, while the leaf blade and sheath proportions were different between two strains. The leaf sheath proportion of BM, representing about 43%, was significantly higher (p<0.01) than that of WT. On contrast, the leaf blade proportion of BM was significantly lower (p<0.01) than that of WT.
Comparison of chemical compositions
Gene mutation greatly altered BM rice straw chemical composition. Compared to WT, the hemicellulose, AIA and CP contents of BM rice straw significantly (p<0.01) increased by 50.3%, 30.6% and 16.9% respectively, while cellulose, ADFom and OM contents significantly (p<0.01) decreased by 33.1%, 29.8% and 4.5% respectively. No significant differences (p>0.05) were detected in ADL and NDFom contents for both strains (Table 1).
SEM and TEM observation
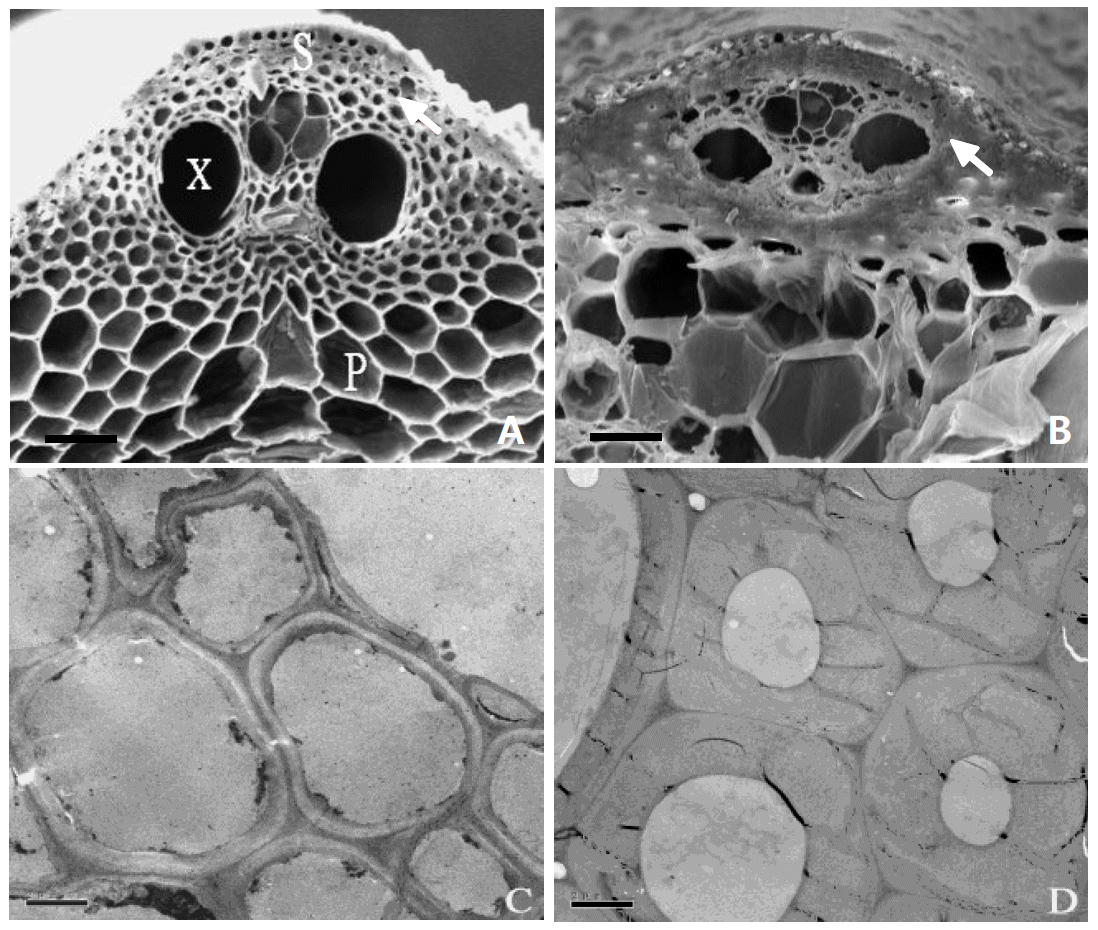
Rice stems are composed of epidermis, sclerenchyma tissue, parenchyma tissue and vascular bundles. BM and WT stems integrally included all the above tissues (Figure 3A and 3B). However, BM had fewer sclerenchyma cells than WT, which are under the epidermal layer (Figure 3A). The alteration in BM sclerenchyma cell walls were identified by TEM observation. Figures 3C and 3D show that the BM had thinner sclerenchyma cell walls than WT.
In situ digestibility
In situ digestibility of BM rice straw was greatly influenced by gene mutation. Table 2 shows that there were no differences in in situ DM disappearance at 24 h of incubation between both strains. However, DM and NDFom disappearance of BM rice straw at 48 h of incubation was significantly greater than WT (40.7% vs 35.5% for DM, 39.7% vs 33.6% for NDFom, p<0.05). For cell wall components, in situ digestibility of cellulose and hemicellulose for BM rice straw was higher than that of WT during incubation time.
As for digestion constants in Table 2, the effective digestibility of DM and NDFom for BM rice straw was significantly higher than WT (31.4% vs 26.7% for DM, 29.1% vs 24.3% for NDFom, p<0.05). Meanwhile, BM had a higher potentially digestible fraction of DM and NDFom and lower lag time than WT. The rate of digestion of the slowly digested fraction of DM and NDFom of BM rice straw was decreased (p<0.05) compared to WT.
DISCUSSION
The mechanical strength of cereal straw is an important factor that influences processing such as chopping, kneading and grinding, which are directly related to the utilization profile of cereal straw. A number of studies have revealed the effects of reducing forage particle size on chewing activities and feeding behavior (Bal et al., 2000; Beauchemin and Yang, 2005). Welch (1982) demonstrated that forage particle size was an important factor in determining the passage of particles from the rumen. Kononoff et al. (2003) revealed that reducing particle size would increase dry matter intake and improve rumen fermentation. In this study, BM rice straw exhibited a fragile culm and leaf (Figure 1). Although the brittle trait could induce lodging during plant growth, it was obvious that this trait was favorable for processing adequate particle size in chewing or feeding.
The chemical composition is an important indicator in the estimation of the nutritional value of feeds and forages. High NDF content could limit rumen intake, and ADF content is negatively related with forage digestibility. In this study BM rice straw contained lower ADFom content and similar NDFom content compared with WT (Table 1). Furthermore, in the grasses, hemicellulose was more digestible than cellulose for ruminants (Keys et al., 1969). This is consistent with our results that in situ digestibility of hemicellulose was higher than cellulose for both strains (Table 2). Agbagla-Dohnani et al. (2001) revealed that rice straw DM degradation was positively related to the hemicellulose content, whereas increasing cellulose and lignin content tended to decrease the DM degradation. BM showed higher hemicellulose content and lower cellulose content than WT, but no difference in lignin content (Table 1). These chemical characteristics imply that BM rice straw could have improved nutritive value.
The plant morphological tissues, including epidermis, sclerenchyma, parenchyma and vascular, had a different response to degradation by ruminal microorganism. SEM and TEM observation showed that sclerenchyma of rice straw stem still remained intact at 72 h incubation, while the parenchyma was digested completely after 48 h of incubation (Wang et al., 2008). For rice straw stem epidermis, most of the cuticle wax layer was intact after 12 h of rumen incubation, while cracking of the cuticle wax and silica layer could be observed after 48 h of incubation (Wang et al., 2007). In this study BM showed fewer sclerenchyma cells and a thinner sclerenchyma cell wall than its wild type (Figure 3). The nutritional value of cereal straws is also related to their botanic characteristics (Abou-El-Enin et al., 1999; Agbagla-Dohnani et al., 2001). Vadiveloo et al. (2000) revealed that the digestibility of rice morphological fractions were different, and followed as: Stem>leaf sheath>leaf blade. BM had larger leaf sheath proportion than that of WT (Figure 2). These morphological and histological characteristics provide an important explanation of why BM rice straw had an improved degradability.
Actually, in most rice-producing countries the improvement of the feeding value of rice straw is out not considered in their modern breeding programs. Brittle culm1 (bc1), which was due to defect in COBRA gene, was the only brittle mutant whose nutritive value has been evaluated (Wang et al., 2006). They showed that there was no difference in organic matter digestibility (OMD) between bc1 and its wild types. However, the DM and NDFom digestibility of BM rice straw were significantly (p<0.01) higher than that of its wild type (Table 2). This inconsistency between BM and Bc1 digestibility could be ascribed to many differences between both, including the mutative gene, plant varieties, and cell wall matrix. Especially, the difference in the cell wall matrix could be the most critical difference, as this determines cellulolytic bacterial adhesion. Further studies on the effects of cellulose synthase gene mutations on the adhesion of rumen cellulolytic bacteria and other rumen microorganism populations may uncover the essential mechanism of improved digestibility of BM rice straw.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







