



INTRODUCTION
Organic rabbit production in Italy must follow the guidelines for organic livestock systems (Council Regulation (EC) No. 1804/1999) and those of two official certification organisms (ICEA, 2007; AIAB, 2012). Only pure breeds, their first generation crosses, and local population are admitted, and red-eye breeds are forbidden. The presence of a large number of animal biodiversity is the basis for the extensive production system, and for this reason it is important to maintain pure breeds, crossed breeds, and local populations that are well suited to pre-existing environmental conditions. Furthermore, the choice of the breed must primarily consider resistance to disease and slow growth, even if reproductive traits and litter performance must be also taken into account. Over the last decade, several authors have studied the effects of alternative and organic rearing systems on the productive live performance, carcass and meat quality of rabbits derived from hybrids, pure breeds or local populations, but only a few have focused on reproductive performance (Cavani et al., 2000, 2004; Dal Bosco et al., 2000, 2002, 2003; Paci et al., 2004, 2005; Pla, 2008; DâAgata et al., 2009; Dalle Zotte et al., 2009; Princz et al., 2009). Many scientific papers and reviews report the effect of season and parity order on reproductive performances of does and productive performances of litters derived from hybrid lines reared under intensive system (Rommers et al., 1999; Marai et al., 2002; Xiccato et al., 2004; Fortun-Lamothe, 2006; Rebollar et al., 2009; Castellini et al., 2010; Villalobos et al., 2010; SzendorĹ et al., 2012), whereas these effects on purebred rabbit does, unselected for high production, and extensively reared are scarce (Paci et al., 2003). The information concerning the effect of the sire genetic origin on litter traits of synthetic lines, native and coloured breeds are also limited (Ozimba and Lukefahr, 1991; Khalil and Al-Saef, 2008; Piles et al., 2008a, b; SzendrĹ et al., 2010, 2012).
The aim of this study was to compare the performance of litters derived from two sire genetic origins, Vienna Blue and Burgundy Fawn, along successive rearing seasons, and doe reproductive performance in an organic production system.
MATERIAL AND METHODS
Animals and housing system
The trial was carried out in a farm certified for organic production located in Lombardia region, Italy (Azienda Agricola âNoi e la Naturaâ, www.noielanatura.it) in 2004 from January to November.
A total of fifty-eight rabbit does at different parity order (P; 1 = nulliparous; 2 = primiparous; âĽ3 = multiparous) and twelve males were involved. The maternal genetic origin was a mixture of crosses of several medium-large size breeds, California and New Zealand White excluded, according to an Italian official certification organism âAssociazione Italiana per lâAgricoltura Biologicaâ (AIAB). The sire genetic origins (SGO) were Vienna Blue (VB; n = 6) and Burgundy Fawn (BF; n = 6) introduced from an outside source at 8 weeks of age.
Does and sires were housed indoor in the same building supplied with a forced-ventilation system and natural lighting. During the experiment, environmental conditions followed seasonality with temperatures ranging from 10°C to 30°C and relative humidity from 60 to 75%. The does were housed individually in wire cages (810Ă60Ă33 cm) with plastic slat floor according to the directives of EC Regulation 1804/1999 and the AIAB (2012). External nest boxes, containing a 2 cm layer of sanitized wood shavings, were provided at 28 d of gestation for kindling and nursing the kits up to weaning. Litters were equalised at birth by cross-fostering (8 to 10 pups per litter).
Management
An organic pelleted diet was given ad libitum to rabbit does and litters during the experimental period. Males were first mated at 5 months of age; nulliparous females were first mated at 3.5 months of age through natural mating. The does were randomly halved and the two groups were assigned to one of the two sire lines for the duration of the experiment.
Does were re-offered to the male 31 to 36 d after each delivery and pregnancy diagnosis through abdominal palpation took place 15 d after each mating. Does diagnosed not pregnant were returned to another buck of the designated sire line for re-mating. The females were re-mated three times until diagnosed as pregnant, after the third negative mating, the females were culled. Along the trial 14 does mated with BF (5 at first, 7 at second, 1 at third and 1 at fourth kindling) and 12 does mated with VB (7 at first, 4 at second and 1 at third kindling) were culled and replaced by other does not previously mated with the males object of study.
The kits, born from January to November, were weaned at 46Âą6 d of age but weaning occurred in two steps: at 35 d of age, the litters were moved to a collective cage in front of the mother that maintained the litter unit before being moved again to the collective cages located in the fattening building. The free suckling system was applied; nest boxes were inspected every morning and dead rabbits were removed.
Reproductive data recording
Doe reproductive performance and the data of 105 litters (55 litters from VB and 50 litters from BF SGO) were recorded throughout the seasons of birth (SB) (winter, spring, summer and autumn). Doe live weight was measured after each delivery. Day of delivery, number of total born, and born-alive per delivery, litter weight of born alive, litter size and weight at 21 d of age, pre-weaning mortality, weaning age, litter size, and weight at weaning were also recorded.
The following data were then calculated: fertility rate (percentage of deliveries/mating), delivery interval and doe LW variation between deliveries, milk output at 21st d of lactation (difference in litter body weight measured before and after suckling at 21 d of age), individual weight of born alive, at 21 d of age, and at weaning, and daily individual growth rate in ranges of 0 to 21 d, 22 d-weaning, and pre-weaning (0-weaning).
Chemical analysis
Throughout the trial, the organic diet was sampled for chemical analysis four times at each batch change (Table 1). AOAC procedures (2000) were used to determine in duplicate dry matter (DM; 934.01), ash (942.05), crude protein (CP; 976.05), crude fibre (CF; 978.10), neutral detergent fibre (NDF; 2,000.04), acid detergent fibre (ADF), and acid detergent lignin (ADL; 973.18) by using the Ankom Technology (Macedon, NY 14502) with filter bags (F57). Ether extract (EE; 920.39) was determined after acid-hydrolysis. Gross energy (GE) was determined by adiabatic bomb calorimeter (ISO, 1998).
Statistical analysis
ANOVA was performed using the SAS (2007) program by including the SGO (VB, BF) the season of birth (SB), the parity (P), and their interactions, for fertility rate only SGO effect was considered. No statistically significant interactions were observed for the considered parameters with exception of SGOxP interaction that resulted statistically significant (p<0.05) only for the variable âindividual weight of born aliveâ.
The statistical significance of the differences was assessed with the Tukey test (SAS, 2007).
RESULTS AND DISCUSSION
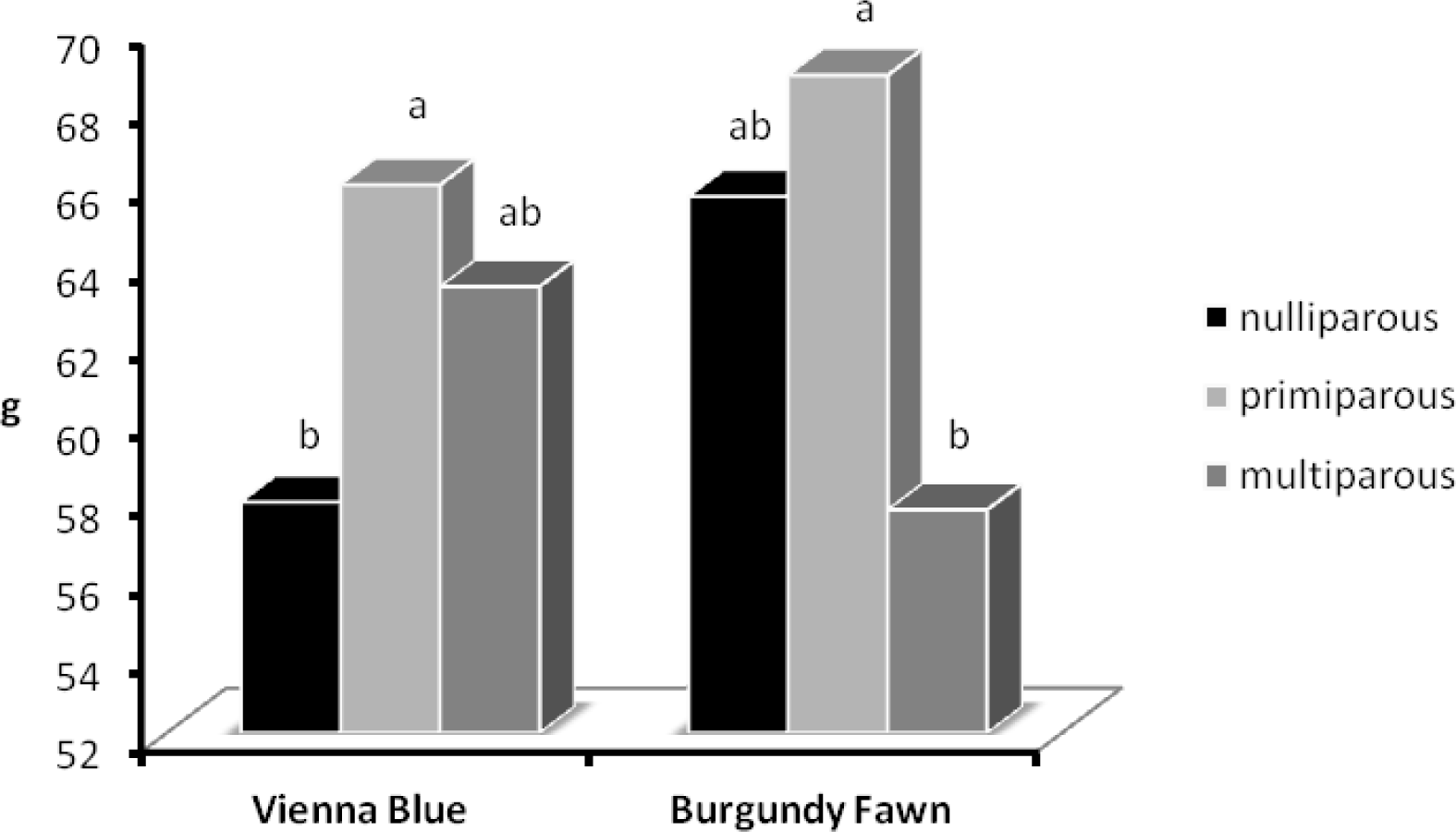
Interaction among main factors was not significant with exception for the SGOxP interaction (p<0.05) for individual weight of born alive pups (Figure 1) and thus Tables do not report the significance of the interaction.
Table 2, 3 and 4 summarize the effects of sire genetic origin (SGO), season of birth (SB), and parity order (P) on doe reproductive performance and on litter size and litter weight at birth, at 21 d, and at weaning, and throughout the 10 months of monitoring.
No significant differences related to the SGO effect were observed for all parameters.
The SB affected almost all the studied parameters. Considering doe LW variations between deliveries (Table 2), a significant LW loss in autumn SB was observed, whereas in winter and spring SB the doe energy balance was positive (â8.18 vs 2.75 and 0.22%, respectively; p<0.05). The slight doe LW loss during summer SB might be explained by the reduced appetite and consequent lower feed intake at moderately high temperatures that induces does to mobilize their body reserves to satisfy their production needs and leads to lower live body weight at kindling (Nardone et al., 2010). The higher doe LW loss during autumn SB could depend on the inability to recover the negative effect of heat suffered during the previous season combined with the energy mobilization required for greater milk production.
Litter size at birth was also affected by SB. The numbers of pups born/delivery and born alive/delivery were higher in winter than in autumn and summer SB (9.5 vs 7.5 and 7.8 pups; 9.4 vs 7.5 and 7.8 pups, respectively; p<0.05, Table 2); similar results were obtained also when considering litter size adjusted by cross-fostering (9.6 vs 7.9 and 7.9 pups; p<0.01, Table 2). The higher litter size constituted by cross-fostering, mainly in winter SB, depended on the decision of the breeder to keep all newborn and equalize the litters to the maximum in order to cover production costs. The SB effect on litter size at birth observed in this study confirms those reported elsewhere (Ayyat and Marai, 1998; Marai et al., 2002; Villalobos et al., 2010; SzendrĹ et al., 2012). The decrease in prolificacy at high environmental temperature depends on a great number of physiological events that may lead to significant reductions in total born and an increase in still-born pups. High environmental temperature has also been shown to compromise oocyte growth and embryo development while increasing embryo mortality (Wolfenson et al., 2000; Marai et al., 2006; Hansen, 2007). The reduced prolificacy observed in autumn may also depend on the effect of the heat stress suffered by the does during the previous hot season. High environmental temperatures are known to lead to reductions in feed intake that impair doe performance and alter the physiological functions that affect reproductive efficiency as a result (Lebas et al., 1986; Lacetera et al., 2003; Fortun-Lamothe, 2006).
SB also affected the individual weight of born alive pups: born alive pups born in winter SB were significantly lighter than those born in autumn SB (58.1 vs 69.1 g; p<0.05, Table 2), probably due to differences in litter size. Birth weight is influenced by several factors during pregnancy, among which, the number of fetuses in the uterine horn plays an important role (Rommers et al., 1999).
Parity order (P) influenced some of the variables considered (Table 2). The LW of multiparous does was significantly higher than that of nulliparous does (4,352 vs 4,122 g; p<0.05). The lighter LW observed in the nulliparous does is related to age: when does start their reproductive life, they have not yet usually achieved total body development (Rebollar et al., 2009). However, the values of LW variation between kindlings indicate an increase of the energy deficit with P despite an increase in doe LW or any extensive reproductive rhythm adopted. Thus, the extensive reproductive rhythm used in accordance with organic farming guidelines does not appear efficient in reducing doe energy deficits, even regardless of parity order. In these females, not selected for high productive and reproductive performances, the after weaning mating was not sufficient to permit a complete recovery of the body reserves lost during lactation. These findings were observed also by other authors in multiparous hybrids, which were unable to cover the requirements for reproduction without mobilizing body energy (Pascual et al., 2000).
The milk output registered at 21st d of lactation (Table 2) was lower in nulliparous and multiparous than in primiparous does (226 and 224 vs 274 g/d; p<0.05). The low milk production of nulliparous does depended on their insufficient feed intake, which led to the consequent inability to cover lactation requirements (Xiccato et al., 2004; Fortun-Lamothe, 2006). The low milk output of multiparous does may also have been related to their body energy deficit, as denoted by the loss of weight and the higher litter size at birth. These results contrast with those reported by Xiccato et al. (2004), who indicated an increase of milk production with increasing P and litter size. It should be noted that the data reported in literature refer to hybrid does, which are selected for very high feed intake, whereas the does used in our research were unselected and for this reason unable to adjust their voluntary feed intake to their energy requirements (Pascual et al., 2000).
As concerns litter performance at birth (Table 2), multiparous doe litters were numerically larger in terms of both total born/delivery (p<0.01) and born alive/delivery (p<0.05), and also heavier (p<0.05) than those of nulliparous and primiparous does (9.4 vs 8.0 and 7.8 pups; 9.1 vs 8.0 and 7.8 pups; 546 vs 479 and 478 g, respectively). In multiparous does, the higher litter weight of born alive was due to the higher number of born alive at kindling and not to pup individual weight, as the latter did not differ significantly among P. Our findings are supported by other recent studies that considered multiparous females inseminated after weaning (Fortun-Lamothe, 2006; Theau-Clement, 2007; Castellini et al., 2010). In our study, does were mated with extensive reproductive rhythm, and therefore the negative effect of lactation on prolificacy was lacking and this led to improved litter parameters.
Data showing the statistically significant (p<0.05) SGOxP interaction for individual weight of born alive pups are reported in Figure 1. In VB SGO, the individual weight of born alive pups slightly increased, whereas in BF SGO it decreased significantly (p<0.05) with P. This individual birth weight decrease (from 68.8 to 57.7 g, in primiparous and multiparous BF, respectively) was probably related to the very high individual weight of pups of part of the does formerly primiparous.
Litter size, litter weight, individual weight, and daily individual growth rate were affected by SB also at 21 d of age (Table 3). In our study, the litter size at 21 d of age showed the lowest values in pups born in summer and autumn compared to those born in winter (6.9 and 7.5 vs 8.8 pups/L, respectively; p<0.01). These results might be explained by the lower number of born alive/delivery observed in summer and autumn and by the slightly higher pre-weaning mortality registered in summer.
In rabbits, the negative consequences of high environmental temperatures are also seen during autumn with the impairment of production and reproduction (Ayyat et al., 1995; Ayyat and Marai, 1998; Marai et al., 2002; Marai and Rashwan, 2004).
SB affected also individual weight at 21st d of age and the daily individual growth rate 0 to 21 d of age, creating heavier rabbits and higher growth rate in rabbits born in summer and in autumn than in the other two previous seasons (387 and 393 g vs 323 and 313 g, p<0.001; 14.9 and 15.3 g/d vs 12.3 and 11.8 g/d, p<0.001; respectively).
Individual weight at 21st d of age is the variable most related to milk output because up to this time the pups consume practically only milk, which contributes to both their growth and development and their survival before weaning (Villalobos et al., 2010). Moreover, individual milk intake is highly dependent on the size of the suckling litter and the milk produced by the doe, which latter is reduced by high environmental temperatures and this reduces doe feed intake in turn.
The best growth of the rabbits born in summer and autumn was ascribed to the significantly lower litter size in these two SB, which provided more milk/pup, and also to the more favourable environmental temperatures during growth, particularly for rabbits born in autumn. On the contrary, the spring SB provided the worst litter performance at 21st d of age because both litters and does had to cope with rising environmental temperatures.
The growth performance of young rabbits at weaning and pre-weaning mortality are summarized in Table 4. Litter size, litter weight, and individual body weight, and daily individual growth rate from 22 d to weaning and during the pre-weaning period were affected only by SB. The litter size of rabbits at weaning was still higher in winter than in summer SB (8.4 vs 6.0 rabbits/litter; p<0.001).
Individual weaning weight was higher in rabbits born in summer than in winter and spring (1,142 vs 1,017 and 971 g, respectively; p<0.05) but the litter weight at weaning was higher in rabbits born in winter due to litter sizes that were higher than those born in spring and summer (8,490 vs 7,240 and 6,789 g/litter; p<0.05), in this way confirming the results found in literature (Ayyat and Marai, 1998; Marai et al., 2002; Marai et al., 2006).
The best individual growth rate during 22 d-weaning was observed in rabbits born in winter; the worst was seen in those born in spring (29.0 vs 23.7 g/d; p<0.05) due to higher solid food intake at lower environmental temperatures.
The individual growth rate of rabbits during the entire pre-weaning period was highest in rabbits born in summer and in winter because they missed the hot season, and was lowest in those born in spring (22.8 and 21.2 vs 19.0 g/d; p<0.01) when young rabbit growth was negatively affected by the heat. This worst rabbit pre-weaning performance observed in the spring SB has also been recently reported by Villalobos et al. (2010).
CONCLUSIONS
After evaluating the reproductive and productive performance of does and their litters it can be concluded that the two sire breeds tested (Vienna Blue and Burgundy Fawn) are comparable and seem to be suitable to the organic production system. The season of birth affected most of the performances of does and young rabbits reared with the organic farming system, and the rabbits seemed better suited to these rearing conditions during winter than in other seasons. The results obtained seem promising: after an initial difficulty in coping with rising temperatures in spring, both does and litters showed a moderately good ability in adapting to adverse environmental conditions, and this was probably due to the genetic origin.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







