




INTRODUCTION
The prevalence rate of obesity has rapidly increased globally in the last two decades, reaching epidemic proportions [1–3]. Numerous studies in both animal models and humans have demonstrated that abnormal adipose tissue accumulation may result in metabolic disorders that cause severe insulin resistance (IR) and type 2 diabetes [4]. Adipose tissue is no longer considered an inert organ for lipid storage and mobilization, but instead thought to play an active role in regulating insulin effects, whole-body energy metabolism, and homeostasis, primarily through its endocrine function [5–8]. As an endocrine organ, adipose tissue secretes a number of adipokines, including leptin and adiponectin, and constitutes a major source of inflammatory cytokines such as tumor necrosis factor alpha (TNF-α) and interleukin-6 (IL-6), mediating obesity-related metabolic dysregulation [9–11]. Most adipokines regulate AMP-activated protein kinase (AMPK) signaling to modulate insulin action [12,13].
AMPK is phosphorylated and activated in response to increased cellular AMP/ATP (adenosine monophosphate/adenosine triphosphate) ratio, thereby acting as an energy sensing enzyme that governs systemic cellular metabolism. It is dysregulated in animals and humans with IR and metabolic syndromes [14,15]. Phosphorylated AMPK promotes glucose uptake into skeletal muscle. Additionally, activated AMPK suppresses gluconeogenesis in the liver and increases fatty acid oxidation in the liver and skeletal muscle. Meanwhile, AMPK inhibits inflammation, endoplasmic reticular stress, and oxidative stress, while inducing autophagy, all of which are involved in the pathogenesis of IR. Interestingly, AMPK is activated by adiponectin in skeletal muscle, leading to induced glucose uptake and improved insulin sensitivity [16]. Adiponectin knockout mice show reduced food intake and decreased AMPK activity. In addition, TNF-α signaling has been shown to suppress skeletal muscle AMPK activity both in vivo and in vitro [17].
However, understanding the link between adipokines and insulin resistance is still distant. The current knowledge of adipokines is based on mouse studies, but subsequent investigations in humans have often produced markedly different results. Therefore, development of animal models that can accurately mimic adipokine action in humans is crucial, and will improve understanding of adipokine mechanisms and their therapeutic effects.
Mini-pig is a well-known animal model used to assess genetic and mechanistic aspects of human obesity and metabolic diseases [18,19]. This study aimed to establish a more appropriate animal model for metabolic diseases to reveal the role of adipokines in human insulin resistance. Therefore, we established and characterized a Bama mini-pig model of obesity, measured plasma glucose and insulin levels, and examined the changes of adiponectin, IL-6 and TNF-α and their association with AMPK activity.
MATERIALS AND METHODS
Study protocols
This study was approved by the Institutional Animal Care and Use Committee of Chinese PLA General Hospital, and were implemented under its supervision (approval document No. 2014-D9-03). Eight Bama mini-pigs with the age of 8 months old and the weight of 25 kg to 26 kg, were fed twice daily a high-fat and high-sucrose diet (55% control diet, 15% beef tallow, and 30% sucrose), for a total of 30 weeks, separately in single cages under controlled environmental conditions. Every 5 weeks, fasting blood glucose (FBG) was measured in all pigs. Intravenous glucose tolerance test (IVGTT) was performed at 30 weeks, and blood samples were collected. The homeostasis model assessment of insulin resistance (HOMA-IR) (fasting plasma glucose×fasting plasma insulin/22.5) was used as an insulin resistance index. According to body weight at 30 weeks, the eight Bama mini-pigs were separated into two groups: obese (n = 3, weight ≥55 kg) and lean (n = 5, weight <55 kg) groups. Three days following IVGTT, skeletal muscle biopsies were obtained under anesthesia and sterile conditions in all pigs.
Intravenous glucose tolerance test
IVGTT was performed at 30 weeks of high-fat and high-sucrose diet (HFHSD). After overnight fasting, baseline blood samples were tested. Pigs were then administered a glucose bolus (500 mg/mL) at a dose of 1 g/kg through a saphenous vein catheter. Blood was collected at indicated time points after glucose administration.
Radioimmunology assays
Blood samples were collected into separation gel vacuum collective tube, and kept on ice until centrifugation (1,500×g, 10 min, 4°C). Serum was then collected and examined. Glucose levels were assessed using a Roche Glucose Kit (HITACHI, cobas8000, Tokyo, Janpan). Three grams adipose tissue of each pig were put into 400 μL lysis buffer in homogenizer to grind into tissue homogenate, and then centrifuged (12,000×g, 15 min, 4°C) twice to get supernatant. Radioimmunology kits (China Institute of Atomic Research, Beijing, China) specific for pig insulin, adiponectin, IL-6, resistin and TNF-α were used. One hundred microliter (μL) serum and adipose tissue supernatant were respectively used for every determination according to the manufacturer’s protocol. Values were calculated as the mean±standard error of the mean (SEM) of duplicate determinations.
Immunoblotting
Tissue lysates were prepared in radioimmunoprecipitation assay (RIPA) buffer containing protease and phosphatase inhibitors. Equal protein amounts were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis, followed by transfer onto polyvinylidene difluoride (PVDF) membranes (Millipore, Darmstadt, Germany). After blocking in 5% non-fat milk in tris-buffered saline (TBS) containing 0.05% Tween 20 (room temperature for 1 h), the membranes were incubated with primary antibodies raised in rabbits against AMPKα (62 kDa), phosphorylated AMPKα-Thr172 (62 kDa), TNF-α (17 kDa), Actin (42 kDa), and Glyceraldehyde 3-phosphate dehydrogenase (GAPDH, 37 kDa) (Cell Signaling Technology, Danvers, MA, USA), respectively. This was followed by incubation with anti-rabbit horseradish peroxidase conjugated secondary antibodies. Signals were visualized using the Super Signal West Pico Chemiluminescent Substrate (Thermo Fisher Scientific, Waltham, MA, USA). Band quantification was performed separately for each gel with the ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
The data are expressed as means±SEM. Statistical significance was determined by unpaired Student’s t-test and two way analysis of variance. Less than 0.05 p value was considered statistically significant. The SPSS software (SPSS, Armonk, NY, USA) was used for statistical analysis.
RESULTS
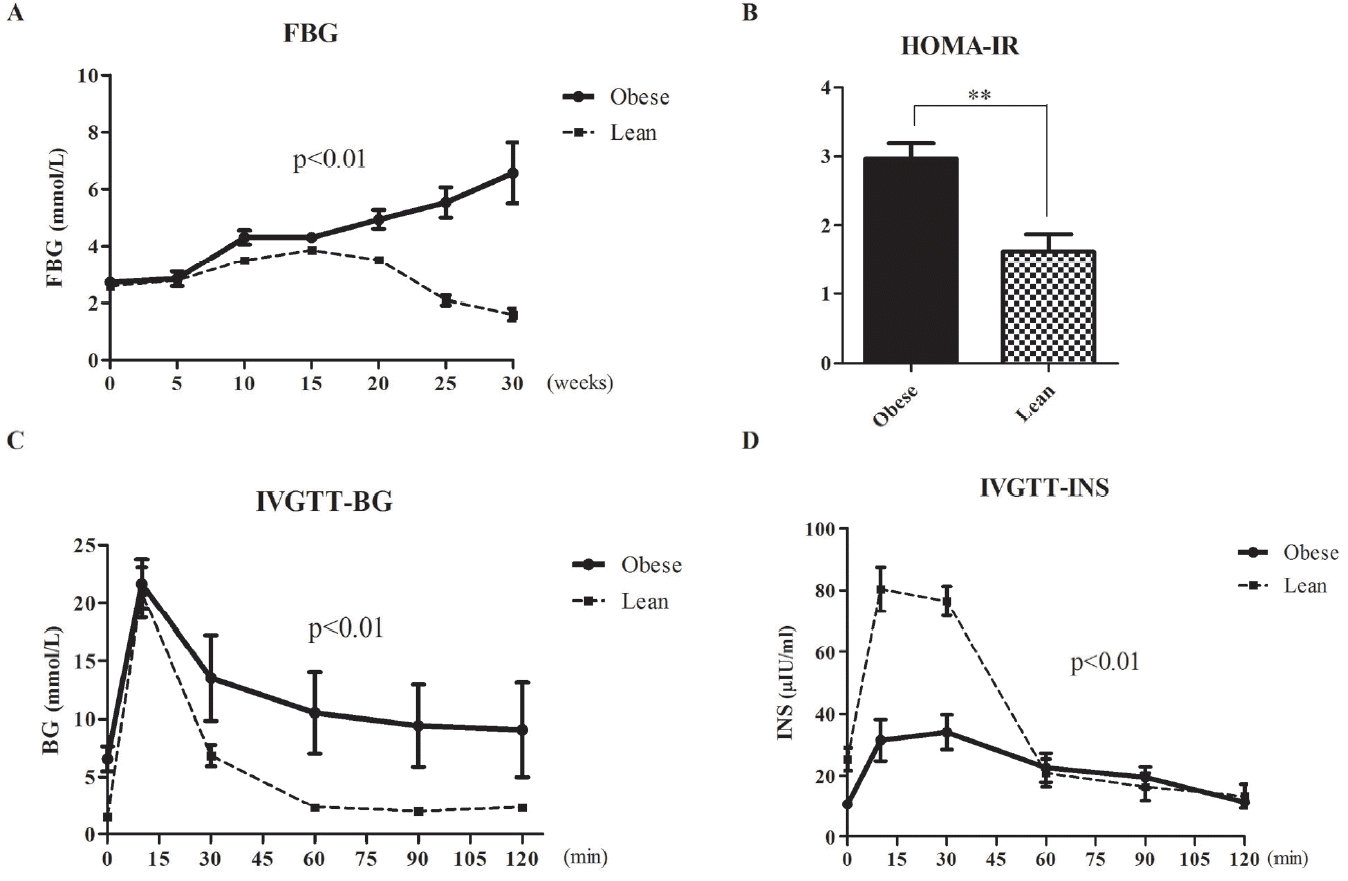
Obese pigs showed hyperglycemia, high triglycerides, and increased HOMA-IR index
As shown in Figure 1A, all animals had similar FBG levels at baseline. During the first 5 weeks, FBG levels were slightly increased in all animals. In obese pigs, they were significantly increased at 10 weeks and maintained increasing till 30 weeks. However, FBG levels in the lean group were beginning to decrease at 20 weeks, and were less than initial value at the end (Figure 1A, p< 0.01). Meanwhile, HOMA-IR levels were significantly increased in obese animals at 30 weeks (Figure 1B, p<0.01). IVGTT results showed that glucose levels were higher in obese pigs compared with the lean group at all time points (p<0.01). In addition, plasma glucose levels in obese pigs remained high for the whole course of IVGTT, while readily returning to normal in lean animals (Figure 1C). In contrast, plasma insulin secretion showed severe deficiency and remained at lower level at almost all time points in the obese group compared with lean animals (Figure 1D, p< 0.01). In obese pigs, total triglyceride and creatinine amounts in serum were elevated compared with their lean counterparts (p< 0.05). However, total cholesterol, low-density lipoprotein, high-density lipoprotein, and serum urea levels had no differences (Tables 1, 2).
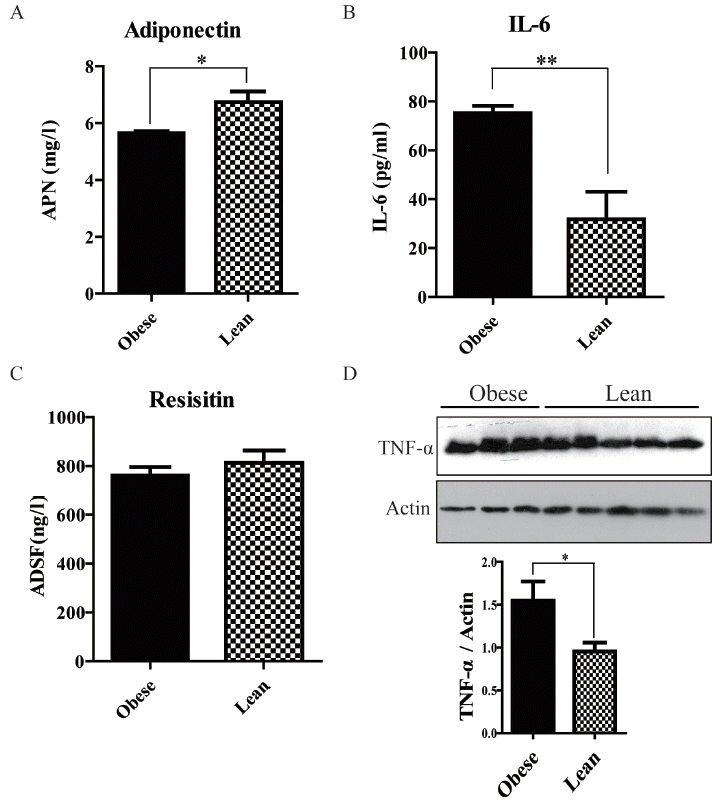
Adipokine levels in obese and lean pigs
As previously reported, multiple adipokines changes are associated with obesity and IR [20–22]. Therefore, we assessed the expression of adiponectin, IL-6, resisitin and TNF-α in adipose tissue and circulating. In serum, adiponectin levels were significantly lower (2.86±1.03 vs 17.50±1.97 ng/mL, p<0.01), while IL-6 amounts were dramatically higher (155.5±19.03 vs 56.28±11.29 pg/mL, p<0.05) in obese animals compared with the lean group (Figure 2A, 2B); meanwhile resisitin (7.64±0.51 vs 6.90±0.24 ng/mL, p>0.05) and TNF-α (19.85±2.42 vs 16.98±0.73 fmol/L, p>0.05) levels showed no difference (Figure 2C, 2D).
In adipose tissue, adiponectin values (5.65±0.07 vs 6.74±0.37 mg/L, p<0.05) were also decreased and IL-6 levels (75.20±2.98 vs 31.80±10.21 pg/mL, p<0.05) were increased in obese pigs than lean group (Figure 3A, 3B). Resisitin levels (760.80±35.10 vs 812.50±50.94 ng/L, p>0.05) still had no difference (Figure 3C). However, the values of TNF-α in adipose tissue were significantly higher in obese pigs (Figure 3D).
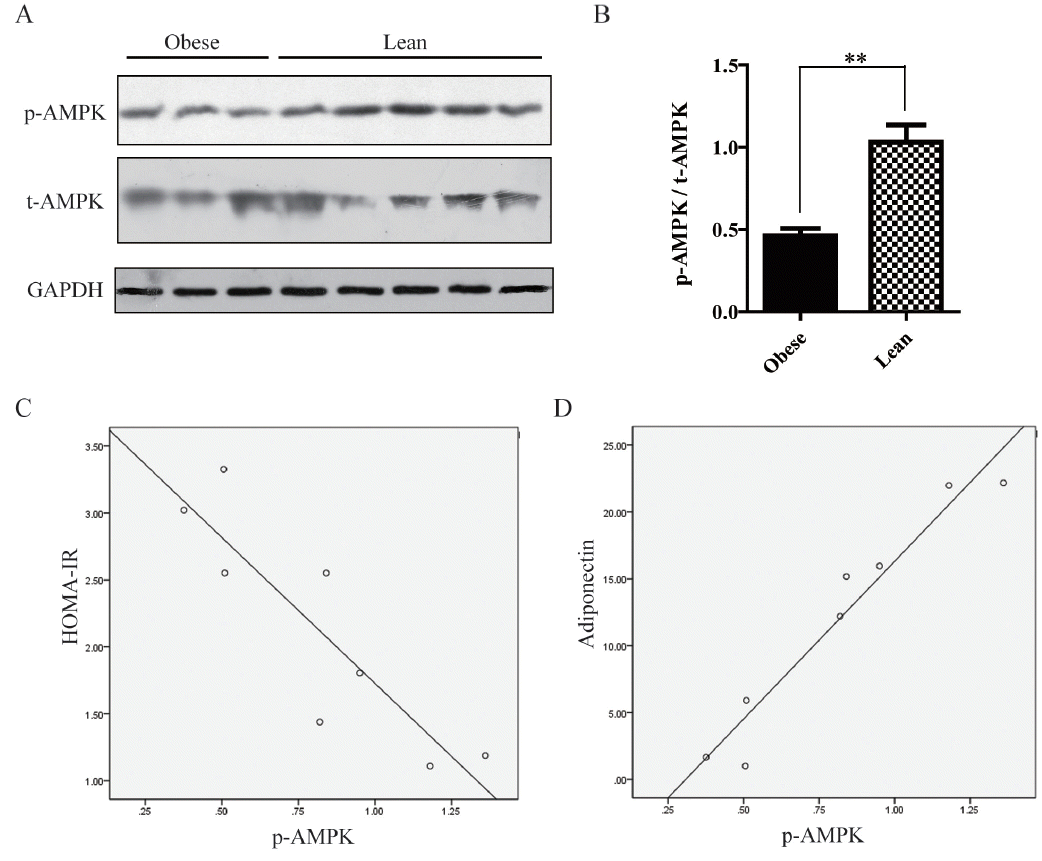
Impairment of AMPK in skeletal muscle
Skeletal muscle insulin resistance could account for over 85% of the whole body impairment in glucose disposal. Therefore, we examined AMPK phosphorylation in skeletal muscle, which is a useful IR marker. Western blot data showed AMPK and GAPDH protein levels were not significantly different between both groups. However, phosphorylated AMPK levels were decreased by 47.4% in obese animals compared to the lean group (Figure 4A, 4B). Pearson correlation analysis showed AMPK phosphorylation was negatively correlated with HOMA-IR index (γs = −0.881, p<0.01), and positively correlated with adiponectin secretion (γs = 0.968, p<0.01), data were shown in Figure 4C and 4D. However, no relationships were found between AMPK phosphorylation and IL-6 amounts (data not shown).
DISCUSSION
Excessive energy intake and a sedentary lifestyle have led to increased incidence of obesity, which is a major risk factor for the development of insulin resistance. This study showed that three of eight Bama mini-pigs fed high-fat and high-sucrose diet for 30 weeks developed obesity. Interestingly, the obese pigs showed hyperglycemia, high triglycerides levels and insulin resistance. With the same HFHSD, FBG levels in obese pigs steadily and rapidly increased over 30 weeks. By comparison, glucose levels slowly and weakly increased in lean pigs only in the first 20 weeks, before gradually returning to normal levels (Figure 1A). In response to the IVGTT, similar to healthy people, insulin release from the pancreas in lean pigs underwent two phases. It is the combination of a robust first phase insulin response followed by a functional second phase insulin response that kept normal blood glucose in lean pigs [23–25]. In contrast, in obese pigs, fasting insulin secretion was severely deficient. Additionally, HOMA-IR levels, which reflect insulin resistance, were significantly increased. Thus, first phase insulin response was too weak to reduce blood glucose in obese pigs. Furthermore, second phase insulin response was also sluggish in these animals, and plasma insulin levels remained low, with only a small peak. Consequently, at the end of the 2 hour IVGTT, blood glucose levels in obese pigs remained high and could not return to normal. Taken together, we have built a new Bama mini-pig model of obesity and impaired glucose tolerance.
As previously reported, multiple adipokines are altered in obesity and play important roles in IR. We proceeded to examine the serum levels of adiponectin, IL-6, resisitin, and TNF-α. Interestingly, obese pigs showed decreased adiponectin and increased IL-6 amounts, as previously reported in obese mice and humans (Figure 2A, 2B, 3A, 3B). Adiponectin, though secreted from adipose tissue, usually decreases in obese state. It is generally admitted that adiponectin are negatively associated with insulin resistance [26]. Adiponectin knockout mice were shown to have decreased AMPK activity and increased susceptibility to insulin resistance; conversely, treatment with adiponectin improves insulin sensitivity. Decreased adiponectin levels are considered a useful predictor of insulin resistance and diabetes in humans. As shown in Figure 4, adiponectin decrease induced AMPK impairment in skeletal muscle, and was strongly associated with IR in obesity Bama mini-pig model. Thus, this model accurately mimicked adiponectin action in humans. Recently, the small molecule adiponectin receptor agonist AdipoRon was discovered, and suggested to be a novel therapeutic target for treating type 2 diabetes, in a mouse model of diabetes [27]. Therefore, further studies in this model will improve understanding of the mechanisms underlying AdipoR agonists for the development of therapeutic interventions.
IL-6 is a pleiotropic cytokine that is secreted by and acts on a wide variety of tissues and cells. Although multiple cells produce IL-6, in healthy humans, adipose tissue releases 10% to 35% of basal circulating IL-6 amounts [28]. Multiple studies showed that release of the adipokine IL-6 is related to obesity; however, conflicting reports exist about the effects of IL-6 on glucose metabolism and insulin sensitivity. Fernandez-Real et al demonstrated a positive association of IL-6 concentration and fasting insulin resistance index in 228 healthy volunteers [29,30]. Subcutaneous injections of recombinant human IL-6 also increases blood glucose levels without changing C-peptide levels, supporting the notion that IL-6 alters insulin sensitivity. Conversely, Carey et al [30] found that infusion of IL-6 accelerated glucose removal with no effects on endogenous glucose production during a hyperinsulinemic-euglycemic clamp study [31]. Similarly, acute infusion of IL-6 was shown to improve insulin sensitivity and glucose removal in animal studies. This discrepancy could be explained by the time course of IL-6 elevation. Chronic elevation of IL-6 levels in obese patients and type 2 diabetics may be associated with insulin resistance, while an acute transient increase can enhance insulin sensitivity. Our data showed obese pigs induced by HFHSD for 30 weeks showed elevated IL-6 levels and insulin resistance, supporting this speculation to a certain extent.
Resistin is a typical example of an adipokine with contrasting roles in rodent and human [32,33]. There are significant differences between these two species with respect to resistin’s gene and protein structure, tissue-specific distribution, and insulin resistance induction [34]. In rodents, resistin is primarily expressed in and secreted from mature white adipocytes [35]. Serum resistin levels positively correlate with body weight in several mouse models of obesity and has been conclusively associated with the development of insulin resistance [36,37]. However, in humans, resistin is majorly derived from peripheral blood mononuclear cells [33]. Although many studies, not all, have reported that plasma resistin was increased in obese individuals, findings of the relationship between circulating resistin levels and obesity and/or insulin resistance in humans remain controversial [32,33, 38]. In this study, we also found that serum resistin levels increased a little in obese pigs, but the values both in serum and adipose tissue showed no statistical difference between obese and lean pigs. Moreover, we failed to identify significant associations between insulin resistance and circulating resistin levels. Our data didn’t support serum resistin being related to insulin resistance in obese pigs.
TNF-α is another example of molecules showing great differences between humans and rodent animals. In obese mice, this cytokine is produced in large amounts by adipocytes and released into the circulation. Insulin resistance can be counteracted by neutralizing TNF-α with specific antibodies. In contrast, human studies revealed that TNF-α is not released into the circulation, but simply constitutes a local factor within the body adipose tissue [39]. Furthermore, intravenous treatment of obese, insulin-resistant humans with a TNF-α antagonist failed to normalize insulin sensitivity [40]. In the model described here, plasma TNF-α levels showed no difference between obese and lean pigs, while the values in adipose tissue were significantly higher in obese pigs. These results were similar with TNF-α changes in obese humans with insulin resistance.
In conclusion, we established and evaluated a novel Bama mini-pig model of obesity, which showed many similarities to human metabolic diseases. This model well established that AMPK impairment induced by adiponectin decrease mediated insulin resistance in HFHSD induction, providing a useful tool for assessing insulin sensitivity and diabetes.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







