




INTRODUCTION
Genetic variability assessment within and among different sheep breeds is essential to develop a successful breeding program for selecting ewes that express superior production traits, e.g. those concerning meat, milk and wool production [1]. Saudi Arabia has three native sheep breeds, namely Najdi, Naeimi, and Herri. The Najdi and Naeimi sheep are fat-tailed, adaptable to prevailing adverse environments and are considered the breeds of choice among Saudi consumers [2]. The Herri sheep breed that has also noticeable adaptability to prevailing adverse environments is an excellent indigenous breed in Saudi Arabia. The genetic determinants of coat color in farm animals, including sheep, are critical for breed recognition and fiber production. Coat color in farm animals is divergent and bears significant biological and economic impacts [3,4]. Melanin is the substance produced by melanocytes that gives skin, hair, and eyes their color. In a large number of mammalian species, the coat color diversity is mainly determined by the relative amount of the two basic melanins, eumelanin (black/brown) and pheomelanin (yellow/red), which are genetically controlled by the extension (E) and agouti (A) loci, respectively [5]. The A locus encodes for the agouti signaling protein (ASIP), a small paracrine signaling molecule [6]. The E locus encodes for the melanocortin-1 receptor (MC1R), a transmembrane protein with seven domains belonging to the G protein-coupled membrane receptors, and localizes on the surface of the melanocyte membrane [7]. When MC1R receptor is activated, it triggers a series of chemical reactions inside melanocytes that stimulate these cells to make eumelanin. If the receptor is not activated or blocked, melanocytes make pheomelanin instead of eumelanin. It has been found that the DNA polymorphisms in MC1R gene may reduce the ability of MC1R to stimulate eumelanin production, causing melanocytes to make mostly pheomelanin [8].
In sheep, the MC1R gene is located on chromosome 14 and has three main alleles, namely E+, Ed, and e, which are defined by three mutations in the coding region and associated with variation in coat color [9–12]. Studies of MC1R gene have provided valuable insights not only into the biology of pigmentation but also the evolution of domesticated animals [13]. So far, there is no report regarding the potential association of MC1R mutations with coat colors in Saudi indigenous sheep. Therefore, the objective of this study was to characterize the DNA polymorphisms of MC1R gene and assess their effects on Saudi Arabian sheep breeds exhibiting different color coats. Individuals of Sawaknee sheep, exotic breed imported from Sudan, were included in this study.
MATERIALS AND METHODS
Samples collection and DNA extraction
To assess MC1R genetic diversity in Saudi sheep, a total of 108 animals were selected from eleven flocks representing three indigenous breeds, namely Najdi (n = 41), Naeimi (n = 36), and Herri (n = 18), in addition to 13 Sawaknee sheep, originally from Sudan. Ten mL of blood samples were collected from each individual sheep by jugular venipuncture into vacuum EDTA tubes. Genomic DNA was extracted using the QIAgen DNeasy blood and tissue DNA extraction kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. The quantity and quality of DNA were checked by Jenway Genova spectrophotometer (Krackeler Scientific Incorporated, Albany, NY, USA). The O.D. 260/280 ratios were between 1.7 and 1.9 indicating high quality DNA [14].
Amplification of MC1R gene
The two primers MF (5′-GAGAGCAAGCACCCTTTCCT-3′) and MR (5′-GAGAGTCCTGTGATTCCCCT-3′) were used to amplify the complete coding region of MC1R gene from the 108 individual sheep [15]. Polymerase chain reaction (PCR) amplifications were carried out in a 25-μL reaction volume containing 100 ng of template DNA and 20 pmol of each primer. To reduce the possibility of cross contamination and variation in the amplification reactions, master mixes containing all PCR reagents including the Kapa Taq polymerase enzyme (KAPA Biosystems, Boston, MA, USA) except DNA template were used. The amplification program was performed using the Gene Amp 9700 thermocycler (Applied Biosystems, Warrington, UK). The amplification protocol was an initial denaturation step for 2 min at 94°C, followed by 35 cycles of 94°C denaturation step for 0.5 min, 60°C annealing step for 0.6 min and 72°C extension step for 1 min. The final step was an extension step at 72°C for 5 min. Electrophoresis of the PCR products was done using 1.5% agarose gel and bands were detected by UV lamp after ethidium bromide staining using gel documentation system (Amersham Biosciences, Uppsala, Sweden).
DNA sequencing and sequence analysis
PCR products of MC1R fragments were cleaned and sequenced by the Advanced Genetic Technologies Center (http://www.uky.edu/Centers/AGTC/). The DNA sequences were edited and aligned using BioEdit software (http://www.mbio.ncsu.edu/Bioedit/bioedit.html). The Basic Local Alignment Search Tool (BLAST) was used to search the GenBank database (http:www.ncbi.nlm.nih.gov) for homologous sequences. The BioEdit software was also used to detect single nucleotide polymorphisms (SNPs) and mutations. MC1R haplotypes and genotypes were determined within and among the studied sheep breeds using BioEdit software.
RESULTS
The amplification of the MC1R gene generated a PCR product of 1,170 bp in length from 41 Najdi, 36 Naeimi, 18 Herri, and 13 Sawaknee sheep. After sequencing, cleaning and aligning the MC1R region from the 108 animals, no SNPs were detected in either the 5′ or 3′ untranslated regions. However, five SNPs were detected in the MC1R complete open reading frame (ORF; 954 bp). Two SNPs were nonsynonymous mutations (c. A218T, p. Met73Lys and c. A361G, p. Asp121Asn), whereas the other three SNPs were synonymous (c. C429T, p. Tyr143Tyr; c. T600G, p. Leu200Leu. and c. C735T, p. Ile245Ile).
Allele frequencies of the 5 SNPs
Each of the five SNPs had two alleles with three different genotypes (Tables 1 and 2). Generally, the allele frequencies of the five positions differed among the studied sheep breeds. Although, all possible alleles of the three synonymous SNPs were detected in the three indigenous Saudi sheep breeds, the Sawaknee breed, imported from Sudan, had only one allele at the five positions (Table 1). In Najdi sheep, the most common allele at position C429T was “T” with a frequency of 91% followed by 0.61% in Herri, while the frequency of the other allele “C” was 0.83% in Naeimi sheep (Table 1). Same pattern was seen in the other two synonymous SNPs (Table 1). The nonsynonymous SNPs, associated with dominant black coat color, at 218 and 361 positions were found in higher frequencies (0.83 and 0.75, respectively) in Najdi breed (generally black with white faces) (Table 1; Figure 1). These two alleles were fixed in Sawaknee sheep (Table 1). In contrast, the Naeimi breed (generally white with brown faces) lacked the A allele at 361 position, where the G allele was fixed (Table 1; Figure 1). The genotype frequencies of the five SNPs varied also among the three Saudi breeds. No heterozygous genotypes of the five SNPs were detected in Sawaknee sheep (generally brown) (Table 2; Figure 1). However, heterozygous genotypes at the five SNP positions were detected in the indigenous Saudi sheep (Table 2). Generally, Najdi sheep showed the highest allelic variation in the MC1R gene, followed by Herri then Naeimi. Sawaknee sheep showed the lowest allelic variation where five out of the 15 expected SNP genotypes were detected (Table 2).
Construction of MC1R haplotypes
The combination of the five mutations in the MC1R gene resulted in eight different haplotypes deposited in GenBank under the accession numbers KU705368-KU705368 (Table 3). The dominant black Ed haplotype H3 (AATGT) was found only in Sawaknee and Najdi sheep (Table 4). The haplotypes H1 (TGCTC), H2 (TGTGT), and H6 (AGTGT) were represented in the three Saudi indigenous breeds. However, the four haplotypes, H4 (TGCGC), H5 (TGCTT), H7 (AGCTC), and H8 (TGTTT), were rare and represented in either one or two breeds (Table 4).
Construction of haplotype combinations
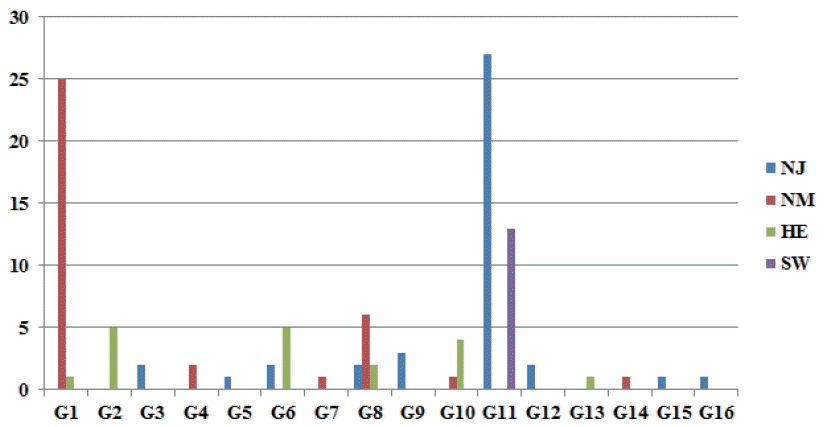
Based on the eight MC1R haplotypes recovered during this study, 36 expected haplotype combinations were constructed. Out of these expected combinations, 16 were observed in the studied sheep. Five out of the 16 MC1R observed haplotype combinations were homozygotes with a total number of 78 individuals. The remaining 11 combinations were represented by 30 animals. Generally, the haplotype combinations were not evenly distributed among the three Saudi breeds (Figure 2). The only haplotype combination that included individual sheep from the three Saudi breeds was the homozygous G8 (H2/H2) (Figure 2). The observed haplotype combinations (G3, G9, G11, and G12) carrying dominant Ed MC1R allele with the nonsynonymous mutations at 218 and 361 positions were mainly from Najdi and Sawaknee sheep (Figure 2). Amongst the 9 Najdi haplotype combinations, 6 were unique whereas 2 out of 7 Herri ones were unique (Figure 2). Out of the 6 Naeimi haplotype combinations, three were unique.
DISCUSSION
The MC1R protein is a highly polymorphic 7-transmembrane G protein-coupled receptor [16]. The MC1R protein is predominantly expressed in melanocytes and plays a central role in regulating melanin production in these specialized cells [8,17]. When the MC1R receptor is bound by its agonist, alpha melanocyte-stimulating hormone (α-MSH), the intracellular levels of cyclic adenosine monophosphate (cAMP) are elevated through a G-protein signaling pathway and the black/brown eumelanin is produced [18]. However, if the MC1R is bound by its antagonist, ASIP expressed by agouti gene, α-MSH binding is blocked, the cAMP levels are reduced and the red/yellow pheomelanin is produced [16,19]. Thus, by varying the levels of α-MSH and agouti protein, the wild type receptor can effectively switch melanin biosynthesis between its two forms [19–21].
In sheep, many loci contributing in coat color, e.g. agouti (A), extension (E), and irregular spotting (S), have been characterized [22,23]. Among these loci, A locus, encoding for ASIP protein, and E locus, encoding for MC1R receptor, are the most studied ones. Although the recessive allele (a) in the A locus can lead to black genotyped sheep, the dominant allele (Ed) in E locus is epistatic over A alleles and produces black genotypes [22,23]. There are other two alleles in the E locus, E+ (wild type) and e [5,12,24,25]. In 1999, Våge et al [9] identified two nonsynonymous mutations that cosegregated with the dominant Ed allele in sheep with black coat colors, the first at position 73 (Met73Lys) and the second at position 121 (Asp121Asn) of MC1R polypeptide. When these two mutations were introduced into mouse MC1R receptors, the Met73Lys mutation showed constitutive activation whereas the Asp121Asn mutation was required for high affinity ligand binding [9]. In this study, these two mutations were detected in Saudi Najdi, generally black, and Sudanese Sawaknee, generally brown, sheep. The Ed haplotype, AATGT, was recovered from either homozygous or heterozygous sheep. Generally, there was a complete association between the presence of Ed allele (AATGT) and black or brown coat color. This haplotype has been previously detected in many black sheep breed worldwide (Table 5). For example, AATGT haplotype was the most common in Chinese Minxian sheep breed expressing black-coat [15]. In addition, this haplotype was observed in Brazilian colored Creole and Italian black Massese sheep breeds [12,25]. Moreover, Ed allele has been observed in other colored sheep breeds including Aragonesa, Romney Marsh black-boned, Kazakh Fat-Rumped, and Xalda [10,15,26].
Although both Met73Lys and Asp121Asn mutations are required for dominant black sheep genotypes, haplotypes having only the Met73Lys mutation, H6 (AGTGT), and H7 (AGCTC), were detected in indigenous Saudi sheep expressing different coat colors. The haplotype H6 was more frequent than H7, in addition occurring in homozygous individuals. These haplotypes were not detected in other sheep breeds worldwide. Våge et al [9] showed that the Met73Lys mutation leads to a constitutive activation of MC1R in sheep. In this study, this mutation led to black coat sheep when it is observed in homozygous individuals, Najdi sheep with genotype H6/H6. However, its occurrence in heterozygous genotypes did not lead to complete black coats, e.g. white Herri with black nose and mouth. It has been reported that single mutations in MC1R gene can lead to either entirely or partially black animals [27–29]. For example, Kijas et al [27] identified a single nonsynonymous mutation at position 121 (Asp121Asn) of MC1R gene in pigs associated with both generally black colored and white or red pigs with black spots. Moreover, Kerje et al [28] showed that the Glu92Lys mutation at chicken MC1R is the causal mutation in black phenotypes of White Leghorn breed.
In addition to the 3 haplotypes of Ed allele, 5 of the wild type E+ allele were detected in Saudi sheep. Out of these 5 haplotypes, 3 (H4, H5, and H8) were rare among the Saudi sheep and only detected in Brazilian sheep [25]. However, the other two common haplotypes (H1 and H2) were detected in other sheep worldwide (Table 5). For example, H1 (TGCTC), the most common haplotype of E+ allele in Saudi sheep, was also the most common haplotype in Chinese Large-tailed Han white breed [15].
The present study supports the suggestion that the two independent nonsynonymous mutations (Met73Lys and Asp121Asn) in MC1R are associated with b pigmentation in sheep. Further studies are carried out to characterize the sequence variations in the lack or red coat colors in Saudi indi-genous Najdi and Sudanese sheep breeds, respectively. Moreover, the Met73Lys mutation was observed in Saudi sheep and it is suggested to be associated with partial black or red pigmentation in sheep. Further studies are carried out to characterize the sequence variations in the agouti locus and their correlation with MC1R variations will be assessed in Saudi sheep breeds.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







