




INTRODUCTION
Corn is a commonly grown crop throughout China and is extensively fed to beef cattle. However, the increasing price of corn, plus the additional subsidy the government is offering farmers to encourage them to grow more corn for the production of ethanol mean that there is growing interest in the use of other crops as an alternative feed source for cattle (Akbar, 2013). Wheat is also a prevalent crop in China, but its nutritive value for ruminants is different from that of corn. Being intermediate in soluble protein, higher in protein and cellulose, and similar in digestible energy and price, wheat is a suitable replacement for corn (Doepel et al., 2009). Studies in beef cattle suggest superior or similar weight gain and feed efficiency when fed wheat vs corn and alfalfa silage. The growth response to wheat silage appears comparable to that with corn silage. Adequate effective fiber with sufficient but not very high fermentable starch, and modest amounts of degradable proteins, make wheat a unique ingredient for sustainable modern ruminant production (Martin et al., 1999). Firkins et al. (2001) reported that total tract dietary of dry matter (DM), organic matter (OM), and neutral detergent fiber (NDF) were not affected by dietary source of carbohydrate; however, total tract acid detergent fiber (ADF) digestibility tended to be lower (p = 0.10) in cows fed the wheat-based total mixed ration (TMR) compared with those fed the corn-based TMR. Nitrogen tended to be lower in cows fed the wheat-based diets compared with cows fed the corn-based diets. Urinary purine derivative excretion was similar in cows fed the corn and wheat-based diets (Gozho et al., 2008).
To explain the responses of beef cattle offered corn and wheat, a more comprehensive understanding of the effects of the diets on ruminal metabolism and digestion is required, but currently only limited data are available. The objectives of this study were to compare cattle fed wheat vs. corn, at three different levels of substitution, on digestibility, digestive enzyme activities, serum metabolite concentrations and ruminal fermentation of beef cattle.
MATERIALS AND METHODS
Study animals
Animals were cared for and handled in accordance with the guidelines for the care and use of laboratory animals of the Animal Nutrition Research Institute of Shandong Agricultural University and the Ministry of Agriculture of China. Four ruminal, proximal duodenum and the terminal ileum cannulated LiLu (Limousin×LuXi) beef cattle, with average live weight of 400±10 kg, were used in this study.
Study design
The four cattle were used in a 4×4 Latin square design experiment and housed in individual pens with free access to fresh water at the farm of Shandong Academy of Agricultural Sciences (Ji’nan, Shandong, China). There were 4 experimental periods with 13 d for dietary adaptation and 3 d for sample collection in each period. During the experimental period, the temperature of the ambient temperature was maintained between 20°C and 28°C.
The treatments comprised a control group (100% corn), 33% wheat group (33% substitution for corn), 67% wheat group (67% substitution for corn), and 100% wheat group (100% substitution for corn) on a DM basis. The experimental diets were designed to achieve the same amounts of crude protein (CP) and metabolizable energy. During the collection period, total fecal and urine collections were carried out for 3 consecutive days. The diets were balanced to be equal in net energy (MJ/kg) and CP (Table 1) and the cattle were fed twice daily, at 0700 and 1700, in equal portions. Supplementary concentrate was offered at 1% per kg body weight (BW), and the concentrate-to-silage ratio was 4:6. The experimental diets met the requirements of all animals for growth and BW gain according to the cattle feeding standard (NY/T815-2004). The wheat and corn were ground using a hammer mill; the geometric mean particle size of the ground wheat and corn was 1.5 mm. The ingredients and chemical composition (mean±standard deviation) of the basal diets (air-dry basis) used to supplement the cattle are shown in Table 1.
Determination of digestibility
The feed intake (kg/d) for each animal was calculated as the difference between the feed offered and that refused during the last 3 d of each period. Total collection of feces and urine was conducted during 3 consecutive days for each period. The excreted feces were collected using pans placed behind the animal and weighed daily. The feces were then thoroughly mixed and one portion was taken to determine the fresh feces nitrogen; a second portion was dried at 55°C for at least 96 h, then sifted through a 40 mesh sieve, and stored at 4°C for determination of DM, OM, ADF, and NDF. The DM was determined by oven drying at 135°C for 2 h (AOAC, 1995; method 930.15). The OM content was calculated as the difference between 100% and the percentage of ash (AOAC, 1995; method 942.05). The NDF was determined as described by Van Soest et al. (1991) using heat-stable α-amylase without sodium sulfite. The ADF was determined according to AOAC (1995; method 973.18). The NDF and ADF values are expressed inclusive of residual ash. For the measurement of CP (N×6.25), samples were ground using a ball mill to a fine powder. Total N was determined by flash combustion and thermal conductivity detection.
Total urine was collected from the bladder of each individual using an indwelling balloon catheter. The urine samples from 3 consecutive days for each period were thoroughly mixed, and 10% 6 mol/L hydrochloric acid at a ratio of 1:50 (acid:urine) was added. About 5% of the volume of this mixture was taken and stored at −20°C for analysis of urinary nitrogen.
Assessment of digestive enzyme activities
The duodenal and ileal digesta were collected through the duodenal and ileal fistulas of each animal on the last day before the morning feed and stored at −80°C for measurement of trypsin, lipase and amylase activity. The digesta were thawed at 4°C, and 8 g portions were added to physiological saline according to a weight/volume ratio of 1:1 at 4°C. The mixture was homogenized for 15 s at low temperatures and then centrifuged at 4°C for 10 min at 1,500×g. The supernatant was collected for measurement of trypsin, lipase and amylase activity by colorimetry (Zhang et al., 2015). The kits were purchased from Nanjing Jiancheng bio Limited (Nanjing, China).
Analysis of blood variables
Blood samples were obtained from the jugular vein into ethylenediaminetetraacetic acid (EDTA, 3 mL) and non-EDTA (10 mL) containing vacutainer tubes before feed delivery on d 0 and at the end of the study, and the complete blood counts recorded. Plasma was then harvested by centrifugation at room temperature for 5 min at 1,500×g and frozen at −20°C for analysis of blood serum metabolites. Serum concentrations of triglycerides, blood urea nitrogen, total cholesterol, high-density lipoprotein, low-density lipoprotein, total protein, blood albumin, aspartate aminotransferase (AST), alanine aminotransferase (ALT) and alkaline phosphatase (ALP) were determined with an Automatic Biochemical Analyzer (7170S, HITACHI, Tokyo, Japan). The concentrations of white blood cells (WBC), lymphocytes, red blood cells, and hemoglobin, and the hematocrit, mean corpuscular volume, mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), platelets, and platecrits were determined using and Automatic Blood Analyzer (KX-21, SYSMEX, Kobe, Japan).
Measurement of ruminal pH and fermentation
Ruminal pH was monitored continuously for 72 h during d 14 to 16 of each experimental period using a Portable PHB-10 pH meter as described by Penner et al. (2006). The daily ruminal pH data were averaged for each 2-h period and used to calculate the mean pH. Ruminal contents were collected on d 15 of each period. Samples were taken at −1, 1, 3, 5, 7, and 9 h after feeding. Approximately 0.5 L of ruminal contents was obtained from multiple sites within the rumen and strained through nylon mesh (pore size 355 μm). Subsamples (5 mL) of filtrate were preserved with 1 mL of 25% (wt/vol) HPO3 and 1 mL of 1% H2SO4 for determination of volatile fatty acid (VFA) and ammonia nitrogen (NH3-N) concentrations, respectively. The samples were stored frozen at −20°C until analyzed. Ruminal VFA were separated and quantified using a gas chromatograph (model 7890A, Agilent Technotogies, Santa Clara, CA, USA) with a capillary column (30 m×0.32 mm i.d., 1 μm phase thickness, Zebron ZB-FAAP, Phenomenex, Torrance, CA, USA). The concentration of NH3-N in the ruminal contents was determined as described by Rhine et al. (1998).
Statistical analyses
The data were analyzed using the MIXED procedures (SAS, Raleigh, NC, USA) for a 4×4 Latin square design. The mixed model included the fixed effects of treatment and the random effects of square, heifer within square, and period within square. Day (or time within day) was considered a repeated measure for variables measured over time. For the repeated measures, various covariance structures were tried, with the final choice depending on low values for the Duncan information criteria. The effect of increasing amounts of wheat in the diet was examined through linear and quadratic orthogonal contrasts using the contrast statement of SAS. Differences were declared significant at p<0.05. Trends were discussed at 0.05≤P <0.10 unless otherwise stated.
RESULTS
Digestibility
The apparent digestibility of DM, OM, and CP was not different among the control, 33% wheat and 67% wheat groups, but greater (p<0.05) for 100% wheat than for control (Table 3). In contrast, the apparent digestibility of NDF and ADF was lower (p<0.05) for 100% wheat than for control. Increased substitution of wheat for corn tended to increase the digestibility of DM and OM as well as that of CP. The digestibility of NDF and ADF decreased as more corn was replaced with wheat.
Digestive enzyme activities
Table 4 showed that increased substitution of wheat for corn significantly increased the activities of trypsin and lipase in the duodenal digesta as well as in the ileal digesta (p<0.05). The activity of amylase in the duodenal digesta increased as more corn was replaced with wheat (p<0.05), but the activity in the ileal digesta did not differ among the groups (p>0.05).
Serum metabolites and whole blood indicators
The hemogram and blood serum chemistry of cattle consuming different amounts of wheat are presented in Tables 5 and 6, respectively. Increased substitution of wheat for corn significantly increased the ALT concentration (p<0.05). There were no significant differences among the groups for the other metabolites (p>0.05). The whole blood indicators did not differ among the groups (p>0.05).
Ruminal pH and fermentation
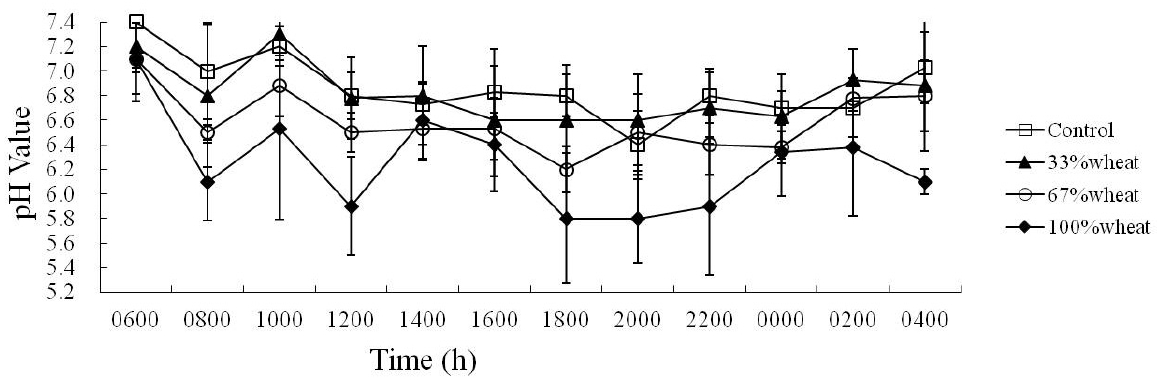
Ruminal pH over 24 h for all the treatments was greatest just before the 0700 h feed; ruminal pH then declined, then showed another peak at 1,000 h, followed by a decline until 2,000 h (Figure 1). This pattern was generally similar among treatments, but the pH in cattle fed 100% wheat diets declined more rapidly than that in cattle fed corn, resulting in a lower ruminal pH between 8,000 and 2,200 h. The ruminal pH was not different between the control and 33%, 67% wheat groups. However, increasing replacement of corn with wheat tended to decrease the mean pH (Table 7).
The contents of individual VFA, such as propionate and butyrate, did not differ between the control and other groups. However, increasing the substitution of wheat for corn increased the molar proportion of acetate and tended to increase the acetate-to-propionate ratio. Cattle fed 100% wheat tended to have the lowest ruminal NH3-N concentration compared with control (p<0.05), whereas no differences were observed among the cattle fed 33% and 67% wheat, compared with control.
DISCUSSION
The attractiveness of wheat is due to its potential for increased yield, increased intake, and ease of preservation compared with other grain sources (GS) (Keady, 2005). Philippeau et al. (1999) reported that the GS (wheat or corn) had little or no effect on DM, OM, or starch intakes. The DM and OM digestibilities in the total tract were greater (p<0.01) for the wheat-based diet than for the corn-based diets. In this study, increasing the content of wheat in the diet resulted in a progressive increase in apparent digestibility of DM, OM, and CP, in agreement with the report by Philippeau et al. (1999). This suggests that increasing the proportion of wheat in the diet may positively affect digestibility, largely reflecting variations in ruminal starch digestion. Starch digestibility was almost complete for all wheat treatments, indicating excellent utilization of the wheat grain, as reported by Sinclair et al. (2003) for wheat of relatively similar chemical composition when using a fecal collection method. We have studied the effects of different methods of processing wheat on nutrient digestibility, digestive enzyme activities and serum metabolite concentrations of beef cattle (Jiang et al., 2015) and found that the digestibility of DM, OM, CP, NDF, and ADF in a puffed wheat diet was significantly higher than that in the other three diets (p<0.05). The digestibility of DM and OM in both the ground wheat diet and the crushed wheat diet was significantly higher than that in squashed wheat diet (p<0.05), and the digestibility of NDF and ADF in the ground wheat diet was significantly lower than that in the other diets (p<0.05). Mean particle size decreases markedly from the cereal to the duodenal contents (i.e., 2,209 vs 416 μm for wheat and 3,182 vs 728 μm for corn), as reported by Mc Geough et al. (2010). Moreover, the starch content in particles decreases markedly for wheat by 45%, for particles >2,000 μm, to 98%, for particles <500 μm, of the initial starch content. In contrast, for corn, the starch content remained almost constant in coarse particles and decreased markedly only for particles <1,000 μm. In this study, the wheat and corn were ground using a hammer mill and the geometric mean particle size of the ground wheat and corn was 1,500 μm. This may explain the lower digestibility of OM, DM, and CP for cattle fed corn diets in the present study.
The lower NDF digestibility of wheat compared with control may be explained by the large fraction of digestible NDF in corn. Nuez-Ortin and Yu (2009) reported greater in situ 48-h disappearance of NDF from corn (45%) vs wheat (51%). Whether the increase in total gastrointestinal tract NDF digestibility that occurs with dietary inclusion of wheat results from increased ruminal or intestinal digestion, or both, is not known. However, the observed ruminal acetate proportion supports the notion of greater ruminal NDF digestion with a wheat diet. The results indicate that substitution of wheat for corn may improve the digestibility of DM, OM and CP, although the digestibility of NDF and ADF may be decreased.
Little information is available on the distribution of starch digestion in the lower digestive tract (small intestine vs hindgut). In contrast to the study of Philippeau et al. (1999), where postruminal digestion of starch was greater in steers fed corn than those fed wheat (34.1% vs 8.8% of starch intake, respectively), our study showed that the activities of trypsin and lipase in the duodenal digesta in steers fed wheat were greater than in those fed corn. Also, the activity of amylase in the duodenal digesta increased linearly (p<0.05) as more corn was replaced with wheat, but the activity in the ileal digesta was not different among groups (p>0.05). The results indicate that, with diets comprising a large amount of starch that escapes ruminal fermentation, a considerable amount of starch could be digested in the hindgut because of limitations on starch digestion in the small intestine. These limitations may involve factors related to the limited time and surface exposure in the small intestine, or additional factors such as a limited capacity of the gut tissue to produce starch-hydrolyzing enzymes (Owens et al., 1986; Harmon, 1993).
Serum concentrations of α-glutamyl transpeptidase, ALT, AST, ALP, and cholesterol are conventionally used to diagnose hepatic damage in humans and domestic animals (Silanikove and Tiomkin, 1992). The α-glutamyl transpeptidase has proved to be a sensitive indicator of minor bovine hepatic damage, whereas ALP and cholesterol are used to detect bile obstruction and mild liver damage (Silanikove and Tiomkin, 1992). In the present study, there were no differences (p>0.10) in the blood serum metabolites or hemogram of the cattle, except for ALT, which increased as wheat was increased in the diet; however, all values were within the normal range for cattle. In addition, there was no difference (p>0.10) in red and WBC counts, indicating that the wheat diet may not have a negative impact on the immune responses of cattle. Lebedeva et al. (2007) found that the activity of the enzyme ALT, which is involved in the integration of protein and carbohydrate metabolism, and its ratio with the activity of AST in the blood of heavy milking Black Pied cows in the dry period characterized the metabolic state of the animals and were associated with their further reproductive capacity. It was shown in the work of Van Kengsel et al. (2007) that the preceding lactation can affect the character and intensity of metabolic processes in the following reproductive cycle in cattle.
Ruminal fermentation characteristics are presented in Table 6 and Figure 1. As expected, the mean pH was less for the wheat-based diet than for the corn-based diets, except for the 33% wheat diet. The pH fell to 5.90 about 5 h after feeding in cattle fed wheat, whereas the minimum pH was 6.70 in cattle fed corn (Figure 1). The pH remained below 6.2 for twice as long in cattle fed wheat compared with cattle fed corn (Philippeau et al., 1999). The fall in pH below 6.2 in relation to the rapid microbial fermentation of starch in wheat may be also involved in the lower fibrolytic activity when wheat replaces corn in the diet (Huhtanen and Khalili, 1992; Martin and Michalet-Doreau, 1995). The increase in the ratio of acetate to propionate as corn was replaced completely resulted primarily from an increase in the molar proportion of acetate and a change in the molar proportion of propionate. Increasing the acetate-to-propionate ration should affect the feed conversion efficiency of growing cattle because less propionate will be provided for gluconeogenesis. The increased molar proportion of acetate suggests that ruminal NDF digestion was increased, possibly due to the reduced ruminal pH.
The ruminal NH3 and plasma urea concentrations observed indicate that adequate amounts of N were available in the experimental diets, with all plasma urea values falling within the range (3.4 to 7.3 mmol/L) defined by Castejon and Leaver (1994) as being normal. The decrease in rumen NH3 observed in response to increasing the content of wheat may reflect an increase in microbial N synthesis, facilitated by the increase in starch intake (Hristov and Ropp, 2003). These results indicate that partial substitution of wheat for corn has a minimal impact on ruminal pH and ruminal fermentability of feed.
IMPLICATIONS
In conclusion, substituting the corn content in feedlot finishing diets with different amounts of wheat improved the digestibility of DM, OM, and CP and increased the activities of digestive enzymes, but reduced the digestibility of NDF and ADF. Serum metabolites and whole blood indicators were not affected by replacement of corn by wheat, except for ALT. These results suggest that the feeding value of a wheat-based feedlot finishing diet is similar to or better than that of a corn-based diet. However, ruminal pH status, the ratio of acetate to propionate and ruminal NH3 were not improved in finishing diets containing 100% wheat as a replacement for corn.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







